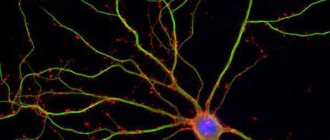
Mouse cortical pyramidal neuron expressing green fluorescent protein (GFP)
Neuron
, or
neuron
(from ancient Greek νεῦρον - fiber, nerve) is a structural and functional unit of the nervous system. A neuron is an electrically excitable cell that processes, stores, and transmits information using electrical and chemical signals. The neuron has a complex structure and narrow specialization. A cell contains a nucleus, a cell body, and processes (dendrites and axons). There are approximately 90–95 billion neurons in the human brain[1][2]. About 4 billion neurons are restored per year. Neurons can connect to one another to form biological neural networks. Neurons are divided into receptor, effector and intercalary.
The complexity and variety of functions of the nervous system are determined by the interactions between neurons. This interaction is a collection of different signals transmitted between neurons or muscles and glands. Signals are emitted and propagated using ions. The ions generate an electrical charge (action potential) that moves through the body of the neuron.
Of great importance for science was the invention of the Golgi method in 1873, which made it possible to stain individual neurons [3][4]. The term “neuron” (German Neuron) to designate nerve cells was introduced by G. W. Waldeyer in 1891 [5] [6].
Structure of neurons
Neuron diagram
Cell body
The body of a nerve cell consists of protoplasm (cytoplasm and nucleus), bounded on the outside by a membrane of lipid bilayer. Lipids consist of hydrophilic heads and hydrophobic tails. The lipids are arranged with hydrophobic tails facing each other, forming a hydrophobic layer. This layer allows only fat-soluble substances (eg oxygen and carbon dioxide) to pass through. There are proteins on the membrane: in the form of globules on the surface, on which growths of polysaccharides (glycocalyx) can be observed, thanks to which the cell perceives external irritation, and integral proteins that penetrate the membrane through, in which ion channels are located.
A neuron consists of a body with a diameter ranging from 3 to 130 microns. The body contains a nucleus (with a large number of nuclear pores) and organelles (including a highly developed rough ER with active ribosomes, the Golgi apparatus), as well as processes. There are two types of processes: dendrites and axons. The neuron has a developed cytoskeleton that penetrates its processes. The cytoskeleton maintains the shape of the cell; its threads serve as “rails” for the transport of organelles and substances packaged in membrane vesicles (for example, neurotransmitters). The cytoskeleton of a neuron consists of fibrils of different diameters: Microtubules (D = 20-30 nm) - consist of the protein tubulin and stretch from the neuron along the axon, right up to the nerve endings. Neurofilaments (D = 10 nm) - together with microtubules, provide intracellular transport of substances. Microfilaments (D = 5 nm) - consist of actin and myosin proteins, especially pronounced in growing nerve processes and in neuroglia. ( Neuroglia
, or simply glia (from ancient Greek νεῦρον - fiber, nerve + γλία - glue), is a collection of auxiliary cells of the nervous tissue. Makes up about 40% of the volume of the central nervous system. The number of glial cells is on average 10-50 times greater than neurons).
A developed synthetic apparatus is revealed in the body of the neuron; the granular ER of the neuron is stained basophilically and is known as the “tigroid”. The tigroid penetrates the initial sections of the dendrites, but is located at a noticeable distance from the beginning of the axon, which serves as a histological sign of the axon. Neurons vary in shape, number of processes, and functions. Depending on the function, sensitive, effector (motor, secretory) and intercalary are distinguished. Sensory neurons perceive stimuli, convert them into nerve impulses and transmit them to the brain. Effector (from Latin effectus - action) - generate and send commands to the working organs. Intercalary neurons - communicate between sensory and motor neurons, participate in information processing and the generation of commands.
There is a distinction between anterograde (away from the body) and retrograde (toward the body) axon transport.
Dendrites and axon
Scheme of the structure of a neuron
Axon is a long process of a neuron. Adapted for carrying excitation and information from the body of a neuron to a neuron or from a neuron to an executive organ. Dendrites are short and highly branched processes of a neuron, which serve as the main site for the formation of excitatory and inhibitory synapses affecting the neuron (different neurons have different ratios of axon and dendrite lengths), and which transmit excitation to the body of the neuron. A neuron may have several dendrites and usually only one axon. One neuron can have connections with many (up to 20 thousand) other neurons.
Dendrites divide dichotomously, while axons give off collaterals. Mitochondria are usually concentrated at branching nodes.
Dendrites do not have a myelin sheath, but axons may have one. The place of generation of excitation in most neurons is the axon hillock - a formation at the point where the axon departs from the body. In all neurons, this zone is called the trigger zone.
Synapse
Synapse
(Greek σύναψις, from συνάπτειν - hug, clasp, shake hands) - the place of contact between two neurons or between a neuron and the effector cell receiving the signal. It serves to transmit a nerve impulse between two cells, and during synaptic transmission the amplitude and frequency of the signal can be adjusted. Some synapses cause depolarization of the neuron and are excitatory, while others cause hyperpolarization and are inhibitory. Typically, stimulation from several excitatory synapses is necessary to excite a neuron.
The term was introduced by the English physiologist Charles Sherrington in 1897.
Brain structure and starting settings
Knowing how a neuron works is not enough to understand what causes rational and intelligent behavior.
Evolution has performed an amazing feat of creating a system from relatively simple elements that can amaze with incredible success in interacting with its environment. It is not enough to take a certain mass of connected neurons (even folded in layers), connect sensors and outputs to it and get at least some semblance of a brain. The main field of work of evolution over millions of years is not the neuron, but the structure and internal organization of nerve cells in the nervous system. In the previous part we talked about the structure of the cerebellar cortex and its example shows that structure and organization is fundamental to its functions. Let's look at how the cerebral cortex is organized, the structure thanks to which Man has become the most successful species on Earth. Table of contents
1. Nervous system simulator. Part 1. Simple adder 2. Nervous system simulator. Part 2. Modulated neuroelement 3. Nervous system simulator. Part 3. The associative neuroelement 4. Memory, memory consolidation and granny neurons 5. Simulation of emotions or the electronic sense of novelty 6. The amazing cerebellum 7. Brain structure and starting settings
It is known that the human cerebral cortex consists of six conventional layers, but it came to this form over many stages of evolution. Therefore, to begin with, we will consider a simplified version with 2 - 3 layers, because this option also occurs in nature. It is more advantageous to study any biological structure from the perspective of its evolutionary development. Evolution followed two paths of development of the cortex: an increase in the number of layers and an increase in the total area of the cortex.
The second feature of the cortex is the presence of so-called cortical columns. We can say that the cortical column is the logical unit of structure of the cortex. The growth of the cortex itself during embryonic development is carried out by division, self-copying of entire columns. And this is logical if we want to maintain some structure while growing.
The cortical columns are based on pyramidal neurons. Pyramidal neurons are one of the largest neurons of the nervous system, and therefore the most studied, since their size allows them to be influenced using special electrodes without destroying the cell. These neurons have a developed system of dendrites, which extends throughout the column, as well as an axon, which in some cases is the source of incoming or outgoing signal. Despite the fact that there may be several such neurons in the first layers of the cortical column, they all work as one neuroelement. The activity of one pyramidal cell often means the activity of the entire group of cells. The evolutionary development of the nervous system was previously focused on reliability, and duplication or distribution of one functionality among groups of cells is a ubiquitous phenomenon. It is impossible to imagine a scheme created by nature in which the death of just one cell would lead to disruption of the functioning of an entire functional unit. We can say that the activity of a group of pyramidal cells in a column indicates the activity of the column itself.
Speakers can be divided into two types: signal generating and signal receiving. The type of column depends on what kind of axons the pyramidal neurons have: afferent or efferent. If there are afferent axons, that is, those bringing a signal, then the column will be activated when a signal is received; the signal may be relayed if there is an efferent axon. In the presence of efferent axons, the result of column activity will be the formation of an outgoing signal.
The dendrites of pyramidal neurons extend to the upper layers of the cortex, where they extend beyond the boundaries of their cortical column, thus causing local interaction between neighboring columns. One of the forms of local interaction is lateral (side) inhibition. Inhibition of neighboring columns is carried out through special inhibitory neurons that are part of the cortical columns. Mainly inhibitory neurons exert their effect on pyramidal neurons, preventing their activation.
Lateral inhibition is exerted on adjacent surrounding columns. It allows you to make the boundaries of the active column areas clearer and the activity areas more localized. Thanks to lateral inhibition, an obstacle to the strong spread of excitation arises.
In addition to lateral inhibition, lateral drive is also present. By adjusting the balance between these two factors, it is possible to very finely regulate the level of overall activity in the brain. For example, sleep requires a reduction in the level of activity; this requires strengthening lateral inhibition and weakening drive. This is done through special chemical signals and mediators.
The effect on neighboring columns does not always take the form of a uniform concentric distribution. The entorhinal cortex is characterized by a distribution that allows arousal to spread more easily in a triangular grid. These are the so-called grid cells, which help animals navigate in space by additionally modifying the image of their place of stay.
The next element of the cortical column is a layer of many relatively small stellate neurons. This layer is usually called granular. The neurons of this layer, due to their size, are less studied than pyramidal neurons. It is the neurons of this layer that can be assigned the main role in the computational processes of the brain; thanks to their work, the formation of associative connections and the formation of images occurs. “Calculations” occur according to the principles of mutual attraction of excitation. Neurons of the granular layer are an associative neuroelement. And the pyramidal cells are assigned the role of a simple adder; these elements seem to separate and highlight images. Pyramidal neurons will be activated when a significant portion of the neurons in the granular layer of the column are excited.
Connections between stellate neurons can freely extend beyond the boundaries of their cortical columns; we can say that the granular layer is almost continuous.
Memory, memory consolidation and granny neurons
Formation of images
Separation of an image by a layer of simple adders
This version of the cortical column and cortical structure is very simple and may even seem primitive, but when used on a large scale and with the right setup, it can produce a structure with high computational performance. Nature has always chosen the simplest, most reliable and effective solutions, and our nervous system is no exception to this rule. I often come across the opinion that a neuron is like some kind of supercomputer or quantum computer that makes complex calculations using some kind of vibrations of ions on its membrane or quantum mechanisms. Even the summation algorithm in the neurons of a conventional perceptron is more complex than in its biological counterpart; in a perceptron, it matters which synapse the signals come from, but in a biological one, only the total amount of impact is important. Stereotypical thinking about the unattainable complexity of the brain can hinder understanding the nature of a biological system.
Surprisingly, the described structure is universal for various types of cortical areas: sensory, motor and associative. There may be variations in the relative thickness of the different layers depending on the function of the cortical area. For example, the motor cortex has an increased layer of pyramidal neurons relative to the granular layer, because the signals from the motor cortex must be clear and strong. And associative areas are characterized by an increased layer of stellate neurons to provide the greatest flexibility in the formation of associative reflex arcs.
Cortical areas are intertwined with many connections, this is done thanks to axons, long processes of neurons. Bundles of axons form nerves, which are the so-called white matter. These nerves can connect both adjacent areas and areas of opposite halves of the brain. Moreover, the architecture of these connections is determined by the evolutionary development of the brain and partially acquired experience and learning, but for different people the picture of these connections will be similar. There are several scientific projects related to mapping these connections, for example, the Human Connectom Project.
Let's look at the principles by which these communications are organized.
The presented diagram is only an example for understanding the principles of the organization. Real circuits in the biological nervous system are many times more complex.
Let's imagine a certain receptor field with a number of receptors of the same type that allow us to obtain certain information about the environment. Certain receptor fields form combined signals, for example, the retina of the eye. Such signals require certain analysis capabilities. Representative columns of these receptors will be distributed throughout the sensory cortex with a certain density while maintaining the topology of the location of the receptors in the receptor field (A). According to the principles of mutual attraction of excitation, certain areas of excitation will form on the cortex, which will be the image of the received combined signal. The primary sensory cortex usually has the highest neuroplasticity, i.e. any combination of excited columns will be processed without taking into account previously received information. The resulting image will be read by other columns, also distributed with a certain density throughout the sensory cortex. These columns will transmit information for further processing to the following areas of the cortex. The nature and density of the “reading” columns provide a certain filter for the resulting images. It is not difficult to understand that this method of processing leads to a significant loss of information received from the receptors; the resulting images do not provide unambiguous information about which receptors were activated. Evolution has chosen two ways to eliminate this problem. Firstly, there is an excess number of receptors, which compensate for the drop in information content after processing. Secondly, if you duplicate information from receptors in another area, but with a violation of the topology of the location of the representative columns in relation to the location of the receptors in the receptor field (B), i.e. Let's confuse them. Then, with many different combinations of receptor activity in two areas, different combinations of images will be formed, which gives more information content and more features that we can identify. Naturally, it should be understood that the “confusion” of signals occurs in a strictly defined way; for example, if you “confuse” the entire field of the retina, this will not give any benefit. Entanglement occurs in small fragments of receptor fields. And of course we can, if necessary, duplicate and confuse the signals more than once. In the nervous system, an example of this division is the dorsal and ventral visual processing pathways.
The general principle of information processing by the cerebral cortex is the sequential transmission of information from area to area with a decrease in the density of connections. Moreover, with each subsequent level, the neuroplasticity of the areas decreases, which adds memory and experience in processing previous information to the calculations. Thus, from the processed information, some key features can be identified that will be associated with specific nerve cells. Simply put, in areas of the cortex with a higher level of processing, the form of excitation distribution that corresponds to the most frequent form of the processed signal will be formed.
The formed image “abc” by a combination of columns a, b and c with frequent repetition or emotional reinforcement will be executed in the future even if the activated combination is incomplete and noisy.
At each level of information processing, a certain time is spent; if it is necessary to analyze the dynamics of changes in information over time, then it is possible to duplicate part of the information from each level into one area. An analogue in biology is the visual information processing area MT (V5), the middle temporal cortex, which collects information from areas V1, V2, V3..., this area is responsible for the perception of movement. When this area is damaged, akinetopsia occurs - the inability to perceive movement.
Associative areas are arranged relatively simply; representations of various sensory and motor areas are located here. Moreover, for motor representations, the presence of bilateral connections is important, either these are axons that can work in both directions, or they are adjacent and jointly working columns at both ends, or two axons of the same column working in different directions. There should be many such areas with different patterns of arrangement of representations, so that the possibility of forming reflex arcs for various combinations is equally possible. Conditioned reflexes are formed in such areas, so these areas should have reduced plasticity.
All connections in the motor and primormotor areas must be bilateral in nature; this is necessary for the formation of associative connections. Essentially, the primormotor area must form reflex arcs of sequences, from one focus of excitation to another. In order for there to be more variability, representations are needed, leading to actions to be repeatedly mixed, this is similar to the situation with sensory processing only in reverse order. Also, all motor actions interact with a specially designed area that coordinates actions in time - the cerebellum.
The presented diagrams are a strong simplification of how it is wired in the brain, and the creation of logical structures similar to the human nervous system is impossible without the participation of specialists in the field of neurobiology and scientists studying the connectome.
What about the other layers?!
– Indeed, I only talked about three layers of the cortex, but in the human brain there are six layers in the cerebral cortex. The cerebral cortex turned out to be a fairly successful product of evolution, even with a small number of layers. The principle of evolution: don’t touch what works. Therefore, all new layers in the cortex are an addition to existing layers. If you look at the layers of the human brain, you can see that we do not have six visible layers, but two logical layers, the structure of which is similar and repeats. Evolution simply repeated existing structures in order to increase productivity.
The pyramidal cells of the outer layer are smaller than the pyramidal cells of the first layer, which means they generally have a higher threshold of sensitivity to activating factors.
The granular layers will operate under similar conditions, but presumably the stellate neurons in the outer layer have lower plasticity, meaning that under some conditions the patterns of activity in the granular layers may differ, despite the fact that the input signals are the same. Thanks to these two logical layers, two modes of activity of the cortical column arise. First: full activity mode, when the pyramidal cells of both layers are activated, the entire column is activated. Second: partial activity mode, when only the upper additional layer is activated. These two operating modes of the speaker can be compared to a person’s ability to speak in a full voice and in a whisper; a whisper is partial activity, and a full voice is full activity.
What does this give? For the sensory cortex, this is an additional level of information processing, as well as the ability to work with images of these areas without activation through receptors. In other words, it gives you the opportunity to work with your imagination. For associative areas, this is an additional level of abstraction, the formation of associations between images that have fewer common features, since the sensitivity threshold of the pyramidal cells of the additional layer is higher. For the motor and primortial cortex, this is an opportunity to work out some movements without directly performing them. Only when the column is fully activated do actions occur; actions with partial activation remain in our imagination.
Of course, there are areas in the brain that control the operation of speaker modes, just as we can very easily change the nature of our speaking from a whisper to full volume. If you increase the level of inhibition in the column, then it is possible that it will only be partially activated; if, on the contrary, you disinhibit the column, then some thoughts can immediately be translated into actions.
Fantasy and a high level of abstract thinking have made humans the most successful species on Earth.
Even if we correctly configure the areas and connections between them, this will not be enough to obtain a working model. Unconditioned reflexes are required. A person is born with a rich set of reflex mechanisms, carefully selected by evolution.
Setting up unconditioned reflexes for the model is an important point, given the fact that learning new reflexes always occurs on the basis of existing reflexes. If any action is not involved in some unconditioned reflex, then it will be impossible to learn to control this action. Biological systems do not initially have “clear” reflexes. After birth, we cannot precisely control our limbs or, for example, walk. This is due to the fact that it is impossible to determine in advance some body parameters, the size of the limbs, their weight, the force created by the muscles, etc. Moreover, these parameters also change dynamically during the growth of the organism. Therefore, many unconditioned reflexes have a certain field of action in the response, and in the head a field of receptors that activate it. An emotional mechanism associated with unconditioned reflexes is also laid down, which will trigger reflexes for correction at a certain point in development.
Let's consider the mechanism for adjusting reflexes using the example of baby babble. In accordance with a certain stage of development, the babbling mechanism is launched, i.e. there is an almost spontaneous launch of “fuzzy” reflexes. When they are started, the child begins to utter various sounds, sometimes these same reflexes are provoked by a sound heard from the outside. The sounds produced often do not correspond to what is expected, i.e. do not correspond to the sounds that trigger the reflex, or trigger hypothetically. The child hears the sounds he himself makes, receiving feedback between the command and the resulting action. Next, the emotional mechanism of novelty comes into force, which connects the center of the need for novelty with a speech act, which gives a new association between the heard sound and the internal motive that prompted the action. Which leads to repeated repetition of the action leading to a saturation of the feeling of novelty. It is claimed that during the period of babbling a child pronounces all the sounds of all languages on Earth. Repeated repetition of sounds leads to the formation of clear patterns of action in accordance with the desired result.
Mastery of the motor-motor system occurs in a similar way. Initially, infants' movements are almost chaotic; there is only an increase in motor activity as a reaction to an emotional stimulus. But over time, a comparison of movements and visual perception, tactile and perception of body position occurs.
Some unconditioned reflexes are not so primitive; in certain cases, template images are embedded in the nervous system, and it is almost impossible to transfer such templates from biological systems to a computer model. Humans have an innate ability to recognize the emotions and movements of individuals of their own species. Therefore, there will be a need to implement some workarounds for some aspects of training.
To bypass the process of long-term training of the motor-motor system through numerous attempts at crawling, standing up, walking and a series of falls, it is possible for android robots to use the method of intercepting control.
A person can transfer his experience of body control to a robot through special devices and motion interception technologies. In a model of the nervous system of a robot with intercepted control, during motor actions the corresponding representations will be activated, as if these movements were performed by the robot itself. Thanks to this, the necessary images and associative connections would be formed. For example, during training with the command: “Raise your hands” - the teacher in the movement interception mode raises his hands, this would lead to the formation of a conditioned reflex between the command and the action, and an associative connection would be formed between the command and the image formed by the processing of position sensors bodies.
In the process of training an electronic model of the brain, it is always possible to control the plasticity of the desired areas, and it is also possible to look “inside” the learning process and highlight, label and strengthen the resulting images. This should significantly speed up the learning process of artificial nervous systems in relation to human learning. As has already become clear, a model built according to the principles described will learn for the most part like a person, without separating learning and interaction with the environment.
Instead of a conclusion
This series of articles ends the next stage of research and development.
The program developed for modeling, although limited in its functionality, made it possible to form a theoretical basis for further work. Of course, the resulting theory still requires further development, as well as practical confirmation and testing. But now it will allow us to take a different look at the problem of modeling the nervous system. The next step is the development of a new modeling environment that will allow us to create models of larger-scale processes occurring in the nervous system. And also to implement a number of ideas, observations and theoretical conclusions. I am not a scientist, and my main activity is not related to the development of intelligent systems, nor is it related to neurobiology.
But I will make every effort to ensure that the project comes to its logical conclusion. I will be glad to consider your suggestions, advice and recommendations, as well as constructive criticism. I will definitely talk about the development news of the project on the pages of GeekTimes and my YouTube channel. Thanks to all! PS It would be inhumane to post the sources without explanation. Therefore, I will post a link to the source and some explanations on the algorithm and logic in a separate article.
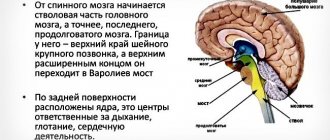
Mechanism of creation and conduction of action potential
In 1937, John Zachary Jr. determined that the squid giant axon could be used to study the electrical properties of axons. Squid axons were chosen because they are much larger than human ones. If you insert an electrode inside the axon, you can measure its membrane potential.
The axon membrane contains voltage-gated ion channels. They allow the axon to generate and conduct electrical signals called action potentials along its body. These signals are generated and propagated due to electrically charged ions of sodium (Na+), potassium (K+), chlorine (Cl-), calcium (Ca2+).
Pressure, stretch, chemical factors, or changes in membrane potential can activate the neuron. This occurs due to the opening of ion channels that allow ions to cross the cell membrane and accordingly change the membrane potential.
Thin axons use less energy and metabolic substances to conduct an action potential, but thick axons allow it to be conducted more quickly.
In order to conduct action potentials more quickly and less energetically, neurons can use special glial cells called oligodendrocytes in the central nervous system or Schwann cells in the peripheral nervous system to cover their axons. These cells do not completely cover the axons, leaving gaps on the axons open to extracellular substance. In these gaps there is an increased density of ion channels. They are called nodes of Ranvier. The action potential passes through them through the electric field between the gaps.
How to choose an activation function?
Now it's time to decide which activation function to use. Should ReLu be used for every case? Or sigmoid? Or tanh? There is no clear answer to these questions. Once you know some characteristics of the function you are trying to approximate, choose an activation function that approximates the function you are looking for better and leads to faster learning.
For example, the sigmoid shows itself well in classification problems (look again at the point about the sigmoid. Does it not have the properties of an ideal classifier?), since the approximation of the classifying function by a combination of sigmoids can be carried out more easily than using ReLu, for example.
Use a function that will make learning and convergence faster . Moreover, you can use your own custom function! If you do not know the nature of the function being examined, then start with ReLu and then work backwards. In most cases ReLu works as a good approximator .
Classification
Structural classification
Based on the number and arrangement of dendrites and axons, neurons are divided into axonless neurons, unipolar neurons, pseudounipolar neurons, bipolar neurons, and multipolar (many dendritic arbors, usually efferent) neurons.
Axonless neurons
- small cells, grouped near the spinal cord in the intervertebral ganglia, which do not have anatomical signs of division of processes into dendrites and axons. All processes of the cell are very similar. The functional purpose of axonless neurons is poorly understood.
Unipolar neurons
- neurons with a single process, present, for example, in the sensory nucleus of the trigeminal nerve in the midbrain. Many morphologists believe that unipolar neurons do not occur in the body of humans and higher vertebrates.
Bipolar neurons
- neurons with one axon and one dendrite, located in specialized sensory organs - the retina, olfactory epithelium and bulb, auditory and vestibular ganglia.
Multipolar neurons
- neurons with one axon and several dendrites. This type of nerve cells predominates in the central nervous system.
Pseudounipolar neurons
- are unique in their kind. One process extends from the body, which immediately divides in a T-shape. This entire single tract is covered with a myelin sheath and is structurally an axon, although along one of the branches the excitation goes not from, but to the body of the neuron. Structurally, dendrites are branches at the end of this (peripheral) process. The trigger zone is the beginning of this branching (that is, it is located outside the cell body). Such neurons are found in the spinal ganglia.
Functional classification
Based on their position in the reflex arc, afferent neurons (sensitive neurons), efferent neurons (some of them are called motor neurons, sometimes this not very accurate name applies to the entire group of efferents) and interneurons (interneurons) are distinguished.
Afferent neurons
(sensitive, sensory, receptor or centripetal). Neurons of this type include primary cells of the sensory organs and pseudounipolar cells, whose dendrites have free endings.
Efferent neurons
(effector, motor, motor or centrifugal). Neurons of this type include the final neurons - ultimatum and penultimate - non-ultimatum.
Association neurons
(interneurons or interneurons) - a group of neurons communicates between efferent and afferent ones.
Secretory neurons
- neurons that secrete highly active substances (neurohormones). They have a well-developed Golgi complex, the axon ends at axovasal synapses.
Morphological classification
The morphological structure of neurons is diverse. Several principles are used to classify neurons:
- take into account the size and shape of the neuron body;
- number and nature of branching of processes;
- axon length and the presence of specialized sheaths.
According to the shape of the cell, neurons can be spherical, granular, stellate, pyramidal, pear-shaped, fusiform, irregular, etc. The size of the neuron body varies from 5 μm in small granular cells to 120-150 μm in giant pyramidal neurons.
Based on the number of processes, the following morphological types of neurons are distinguished[7]:
- unipolar (with one process) neurocytes, present, for example, in the sensory nucleus of the trigeminal nerve in the midbrain;
- pseudounipolar cells grouped near the spinal cord in the intervertebral ganglia;
- bipolar neurons (have one axon and one dendrite), located in specialized sensory organs - the retina, olfactory epithelium and bulb, auditory and vestibular ganglia;
- multipolar neurons (have one axon and several dendrites), predominant in the central nervous system.
Functions of motor neurons
Afferent neurons are part of the peripheral nervous system.
Their perceiving sections - receptors - are located in the skin, the walls of tubular organs or blood vessels, and the capsules of parenchymal organs. Afferent neurons are pseudounipolar. That is, they have one process that extends from the body and then divides into two. Thus, one process approaches the body from the receptor, and then another extends from it to the center.
The body itself is located in the spinal ganglia, or nodes. These formations consist entirely of nerve cell bodies.
The main function of the afferent neuron has already been noted above - the perception of information from the outside using a receptor. But what does this phrase mean?
The perception of any signal is based on the process of information analysis. Its essence lies in the detailed “decomposition” of the stimulus into individual components, careful study and identification of its individual properties. But only a superficial analysis of information occurs in the receptor. Therefore, one reflex arc is not capable of providing complex reactions. For example, one of the reflexes is extension of the knee to a blow to the tendon with a hammer.
A more subtle “decomposition” of the stimulus is carried out in the central nervous system. And this process is ensured most subtly and thoroughly by the cerebral cortex as the most historically new structure of the central nervous system.
Thus, the afferent neuron is an important part of the reflex arc, thanks to which we are able to perceive external irritation and quickly respond to it.
All physical actions that a person can perform are carried out according to the same principle: through contraction and stretching of muscles and tendons. These contractions occur due to the existence of communication between all muscles and tendons with a single coordination center - the brain. These messages consist of cells with different tasks - neurons. Accordingly, special motor cells—motoneurons—participate in the implementation of motor functions.
Muscle contraction occurs by changing just two commands: relax and tense - that is, straighten and contract. A special motor neuron is responsible for each of these conditions. The motor neuron responsible for contraction is called a flexor, and the motor neuron responsible for relaxation is called an extensor.
All muscle fibers have their own motor neurons attached. Together, the motor cell and the muscle fiber to which it is attached are called a “motor unit.” Each such unit functions independently of other similar units. And each motor unit contains only one type of muscle fiber.
Types of muscle fibers:
- Slow oxidative fibers.
- Fast oxidative fibers.
- Fast glycolytic fibers.
Features of nerve cells
Neurons are somewhat reminiscent of a colony of ants - there are just as many of them and they are divided into various groups according to specialization. It is in the differences between these specializations that their specific features and differences lie.
Types of motor neurons, their characteristics and localization in the cerebral cortex:
- Central innervating flexors: localized in the area of the precentral gyrus and are responsible for compression (contraction) of skeletal muscles.
- Central innervating extensors: localized in the hindbrain region and are responsible for relaxing skeletal muscles.
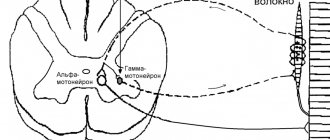
- Peripheral alpha: cells that transmit commands to muscle fibers to contract. Localized in the anterior horns of the spinal cord.
- Peripheral gamma: cells responsible for muscle tone. They are localized in the same place, in the anterior horns of the spinal cord.
- Intercalary: present in all parts of the central nervous system, and carry out the role of communication of all signals in the central nervous system.
Considering that neurons ensure the functioning of all body systems, the functions of nerve cells must be very diverse. Moreover, all of them have not yet been fully clarified. Among the many different classifications of these functions, we will choose one that is the most understandable and closest to the problems of psychological science.
This is the main function of neurons, with which others are associated, although no less significant. This same function is also the most studied. All external signals received by the organs enter the brain, where they are processed. And then, as a result of feedback in the form of impulses-commands, they are transferred along the efferent nerve fibers back to the sense organs, muscles, etc.
This constant circulation of information occurs not only at the level of the peripheral nervous system, but also in the brain. The connections between neurons exchanging information form incredibly complex neural networks. Just imagine: there are at least 30 billion neurons in the brain, and each of them can have up to 10 thousand connections.
Neurons are responsible for what we call memory. More precisely, as neurophysiologists have found, the preservation of traces of signals passing through neural circuits is a kind of side effect of brain activity. The basis of memory is the same protein molecules - neurotransmitters, which arise as connecting bridges between nerve cells.
This ensures interaction between different parts of the brain. Instant “flares” of transmitted and received signals, foci of increased excitation in the cerebral cortex - this is the birth of images, feelings and thoughts. Complex neural connections that connect different parts of the cerebral cortex and penetrate into the subcortical zone are the product of our mental activity. And the more such connections arise, the better the memory and the more productive the thinking. That is, in essence, the more we think, the smarter we become.
The activity of nerve cells is not limited to information processes. Neurons are real protein factories. These are the same neurotransmitters that not only act as a “bridge” between neurons, but also play a huge role in regulating the functioning of our body as a whole. Currently, there are about 80 types of these protein compounds that perform various functions:
- Norepinephrine, sometimes called the rage or stress hormone. It tones the body, increases efficiency, makes the heart beat faster and prepares the body for immediate action to repel danger.
- Dopamine is the main tonic of our body. It is involved in the activation of all systems, including during awakening, during physical activity, and creates a positive emotional mood, even euphoria.
- Serotonin is also a “good mood” substance, although it does not affect physical activity.
- Glutamate is a transmitter necessary for memory function; without it, long-term storage of information is impossible.
- Acetylcholine controls the processes of sleep and awakening, and is also necessary for enhancing attention.
Neurotransmitters, or more precisely their quantity, affect the health of the body. And if there are any problems with the production of these protein molecules, then serious diseases can develop. For example, a lack of dopamine is one of the causes of Parkinson's disease, and if too much of this substance is produced, schizophrenia can develop. If not enough acetylcholine is produced, then very unpleasant Alzheimer's disease can occur, which is accompanied by dementia.
The formation of brain neurons begins even before a person is born, and throughout the entire period of growing up, the active formation and complication of neural connections occurs. For a long time it was believed that new nerve cells cannot appear in an adult, but the process of their death is inevitable.
But recent studies have refuted this pessimistic forecast. Swiss scientists have proven that there is a part of the brain that is responsible for the birth of new neurons. This is the hippocampus; it produces up to 1,400 new nerve cells every day. And we can only more actively include them in the work of the brain, receive and comprehend new information, thereby creating new neural connections and complicating the neural network.
Neuron development and growth
Growth cone
The issue of neuronal division currently remains controversial. According to one version, a neuron develops from a small precursor cell, which stops dividing even before it releases its processes. The axon begins to grow first, and dendrites form later. At the end of the developing process of the nerve cell, a thickening appears, which makes a path through the surrounding tissue. This thickening is called the growth cone of the nerve cell. It consists of a flattened part of the nerve cell process with many thin spines. The microspinuses are 0.1 to 0.2 µm thick and can reach 50 µm in length; the wide and flat region of the growth cone is about 5 µm in width and length, although its shape can vary. The spaces between the microspines of the growth cone are covered with a folded membrane. Microspikes are in constant motion - some are retracted into the growth cone, others elongate, deviate in different directions, touch the substrate and can stick to it.
The growth cone is filled with small, sometimes connected to each other, membrane vesicles of irregular shape. Under the folded areas of the membrane and in the spines there is a dense mass of entangled actin filaments. The growth cone also contains mitochondria, microtubules and neurofilaments, similar to those found in the body of the neuron.
Microtubules and neurofilaments elongate mainly due to the addition of newly synthesized subunits at the base of the neuron process. They move at a speed of about a millimeter per day, which corresponds to the speed of slow axonal transport in a mature neuron. Since the average speed of advancement of the growth cone is approximately the same, it is possible that during the growth of the neuron process, neither the assembly nor destruction of microtubules and neurofilaments occurs at its far end. New membrane material is added at the end. The growth cone is an area of rapid exocytosis and endocytosis, as evidenced by the many vesicles present there. Small membrane vesicles are transported along the neuron process from the cell body to the growth cone with a stream of fast axonal transport. Membrane material is synthesized in the body of the neuron, transported to the growth cone in the form of vesicles and incorporated here into the plasma membrane by exocytosis, thus lengthening the process of the nerve cell.
The growth of axons and dendrites is usually preceded by a phase of neuronal migration, when immature neurons disperse and find a permanent home.
Sigmoid
The sigmoid appears smooth and similar to a step function. Let's consider its advantages.
Firstly, the sigmoid is nonlinear in nature, and the combination of such functions also produces a nonlinear function. Now we can stack layers.
Another advantage of such a function is that it is not binary, which makes the activation analog, unlike a step function. The sigmoid is also characterized by a smooth gradient.
If you notice, in the range of X values from -2 to 2, the Y values change very quickly . This means that any small change in the value of X in this region entails a significant change in the value of Y. This behavior of the function indicates that Y tends to be pushed towards one of the edges of the curve.
The sigmoid does seem to be a suitable function for classification problems. It strives to bring the values to one of the sides of the curve (for example, to the upper one for x=2 and the lower one for x=-2). This behavior makes it possible to find clear boundaries when making predictions.
Another advantage of the sigmoid over the linear function is the following. In the first case, we have a fixed range of function values - [0,1], while the linear function varies within (-inf, inf). This property of the sigmoid is very useful, since it does not lead to errors in the case of large activation values.
Today, the sigmoid is one of the most common activation functions in neural networks. But it also has disadvantages that are worth paying attention to.
You may have already noticed that as you approach the ends of the sigmoid, the Y values tend to respond little to changes in X. This means that the gradient in such areas takes on small values. This, in turn, leads to problems with the extinction gradient. Let us consider in detail what happens as the activation function approaches the almost horizontal part of the curve on both sides.
In such a case, the gradient value is small or disappears (cannot make a significant change due to the extremely small value). The neural network refuses to learn further or does so extremely slowly (depending on the method of use or until the gradient/computation begins to suffer from floating point limitations). There are options for working on these problems, and the sigmoid is still very popular for classification problems.
Properties and functions of neurons
Properties:
- The presence of a transmembrane potential difference
(up to 90 mV), the outer surface is electropositive with respect to the inner surface. - Very sensitive
to certain chemicals and electrical current. - The ability to neurosecretion
, that is, to synthesize and release special substances (neurotransmitters) into the environment or synaptic cleft.
- High energy consumption
, high level of energy processes, which necessitates a constant influx of main energy sources - glucose and oxygen, necessary for oxidation.
Functions:
- Receiving function
(synapses are contact points; we receive information in the form of an impulse from receptors and neurons). - Integrative function
(information processing, as a result, a signal is generated at the neuron output that carries information from all summed signals). - Conductor function
(information flows from the neuron along the axon in the form of electric current to the synapse). - Transmitting function
(a nerve impulse, having reached the end of an axon, which is already part of the structure of the synapse, causes the release of a mediator - a direct transmitter of excitation to another neuron or executive organ).
Hyperbolic tangent
Another commonly used activation function is the hyperbolic tangent.
The hyperbolic tangent is very similar to the sigmoid. Indeed, this is an adjusted sigmoid function.
Therefore, such a function has the same characteristics as the sigmoid discussed earlier. Its nature is non-linear, it is well suited for layer combinations, and the range of function values is (-1, 1). Therefore, there is no point in worrying that the activation function will be overloaded with large values. However, it is worth noting that the gradient of the tangential function is greater than that of the sigmoid (the derivative is steeper). The decision of whether to choose sigmoid or tangent depends on your gradient amplitude requirements. Just like the sigmoid, the hyperbolic tangent has the problem of gradient disappearance.
Tangent is also a very popular and used activation function.
Notes
- Williams RW, Herrup K.
The control of neuron number. (English) // Annual review of neuroscience. - 1988. - Vol. 11. - P. 423-453. — DOI:10.1146/annurev.ne.11.030188.002231. - PMID 3284447. [correct] - Azevedo FA, Carvalho LR, Grinberg LT, Farfel JM, Ferretti RE, Leite RE, Jacob Filho W., Lent R., Herculano-Houzel S.
Equal numbers of neuronal and nonneuronal cells make the human brain an isometrically scaled-up primate brain . (English) // The Journal of comparative neurology. - 2009. - Vol. 513, no. 5. - P. 532-541. — DOI:10.1002/cne.21974. - PMID 19226510. [correct] - Camillo Golgi (1873). “Sulla struttura della sostanza grigia del cervelo.” Gazzetta Medica Italiana. Lombardia 33
: 244-246. - Isaac Asimov.
A Brief History of Biology. - Ripol Classic, 2013. - P. 114. - Jean-Pierre Changeux, Laurence Garey.
Neuronal Man – The Biology of Mind. - Princeton University Press, 1997. - P. 28. - Namely, the term "neuron" appears in the quotation "Das Nervensystem besteht aus zahlreichen, untereinander anatomisch wie genetisch nicht zusammenhängenden Nerveneinheiten (Neuronen)" from the following source: Heinrich Wilhelm Gottfried von Waldeyer-Hartz (1891). “Ueber einige neuere Forschungen im Gebiete der Anatomie des Centralnervensystems.” Deutsche medicinische Wochenschrift 17
(50): 1352. DOI:10.1055/s-0029-1206907. - cm.
Types of neurons
The described cells are classified according to functionality and structural features. In the first case, sensitive, intercalary, and motor neurons are distinguished. Alternative medical names are afferent, association, and command-efferent. Secretory neurons are considered separately. These cells produce hormones, highly active substances involved in the correction of endocrine balance, regulating emotions and psychological background. The structure of a neuron suggests the following classification:
- unipolar
- have only one process; - bipolar
- equipped with a dendrite on one side and an axon on the other side; - pseudounipolar
- one process, but it branches; - multipolar
- the most common, having many “scattered” dendrites. Axon in one copy.
Literature
| Neuron at Wikimedia Commons |
- Polyakov G.I., On the principles of neural organization of the brain, M: Moscow State University, 1965
- Kositsyn N. S. Microstructure of dendrites and axodendritic connections in the central nervous system. M.: Nauka, 1976, 197 p.
- Nemechek S. et al. Introduction to neurobiology, Avicennum: Prague, 1978, 400 p.
- Bloom F., Leiserson A., Hofstadter L. Brain, mind and behavior
- Brain (collection of articles: D. Hubel, C. Stevens, E. Kandel, etc. - issue of Scientific American (September 1979)). M.:Mir, 1980
- Savelyeva-Novoselova N. A., Savelyev A. V. Device for modeling a neuron. A. s. No. 1436720, 1988
- Savelyev A.V.
Sources of variations in the dynamic properties of the nervous system at the synaptic level // Journal “Artificial Intelligence”, NAS of Ukraine. - Donetsk, Ukraine, 2006. - No. 4. - P. 323-338.
What is a neuron?
The organic unit shown represents the basic element of the brain. There are many terms for what neurons are, but the general definition is a cell that is subject to electrical and biochemical excitability. It is intended for receiving, transmitting and issuing information, its analysis and accumulation. The structure of a neuron suggests its relationship with other similar cells. Thanks to this, highly functional nerve networks are formed.