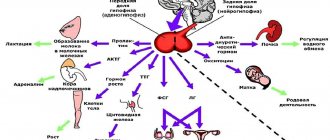
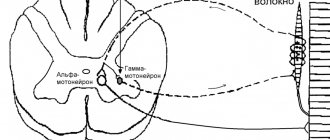
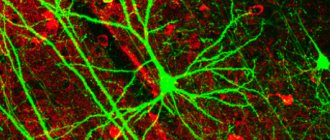
For the term "fiber" see also other meanings.
Nerve fibers
- long processes of neurons covered with glial membranes. Nerve impulses travel along nerve fibers, along each fiber separately, without reaching others [1] [2].
In different parts of the nervous system, the sheaths of nerve fibers differ significantly in their structure, which underlies the division of all fibers into myelin
and
unmyelinated
.
Both consist of a nerve cell process lying in the center of the fiber, and therefore called the axial cylinder
(axon), and, in the case of myelin fibers, a surrounding sheath. Depending on the intensity of the functional load, neurons form one or another type of fiber. The somatic part of the nervous system, which innervates the skeletal muscles, which has a high degree of functional load, is characterized by a myelin (pulpless) type of nerve fibers, and the autonomic part, which innervates the internal organs, is characterized by a non-myelinated (pulpless) type.
Sheathed plexuses of bundles of nerve fibers form nerves.
Erlanger-Gasser classification[ | ]
It is the most complete classification of nerve fibers based on the speed of nerve impulse conduction.
| Fiber type | Function | Diameter, microns | Conduction speed, m/s | Myelination |
| Aα | Afferent - muscle spindles, tendon organs; efferent - skeletal muscles | 10-20 | 60-120 | + |
| Aβ | Afferent - tactile sense; collaterals of Aα fibers to intrafusal muscle fibers | 7-15 | 40-90 | + |
| Aγ | Efferent - muscle spindles | 4-8 | 15-30 | + |
| Aδ | Afferent - temperature, rapid pain transmission | 3-5 | 5-25 | + |
| B | Sympathetic, preganglionic; postganglionic fibers of the ciliary ganglion | 1-3 | 3-15 | intermittent |
| C | Sympathetic, postganglionic; afferent - slow pain conduction | 0,3-1 | 0,5-2 | — |
Structure and functions of nerve fibers, their classification
Nerve fibers are processes of nerve cells (neurons) that have a membrane and are capable of conducting nerve impulses.
The main component of the nerve fiber is the process of the neuron, which forms, as it were, the axis of the fiber. Mostly this is an axon. The nerve process is surrounded by a membrane of complex structure, together with which it forms a fiber.
Nerve fibers are divided into myelinated and unmyelinated. The former have a myelin sheath covering the axon, the latter lack a myelin sheath.
Myelin fibers predominate in both the peripheral and central nervous systems. Nerve fibers lacking myelin are located predominantly in the sympathetic division of the autonomic nervous system.
Structure of nerve fibers:
The myelinated nerve fiber contains the following elements (structures): 1) an axial cylinder located in the very center of the nerve fiber,
2) the myelin sheath covering the axial cylinder,
3) Schwann shell.
The axial cylinder consists of neurofibrils. The pulpy membrane contains a large amount of lipoid substances known as myelin. Myelin ensures the speed of nerve impulses. The myelin sheath does not cover the entire axial cylinder, forming gaps called nodes of Ranvier. In the area of the nodes of Ranvier, the axial cylinder of the nerve fiber is adjacent to the superior Schwann membrane.
The fiber space located between two nodes of Ranvier is called a fiber segment. In each such segment, the nucleus of the Schwann membrane can be seen on stained preparations. It lies approximately in the middle of the segment and is surrounded by the protoplasm of the Schwann cell, the loops of which contain myelin. Between the nodes of Ranvier, the myelin sheath is also not continuous. In its thickness, so-called Schmidt-Lanterman notches are found, running in an oblique direction.
Schwann membrane cells, as well as neurons with processes, develop from the ectoderm. They cover the axial cylinder of the nerve fiber of the peripheral nervous system, similar to how glial cells cover the nerve fiber in the central nervous system. As a result, they may be called peripheral glial cells.
In the central nervous system, nerve fibers do not have Schwann sheaths. The role of Schwann cells here is performed by elements of oligodendroglia. An unmyelinated (unmyelinated) nerve fiber is devoid of a myelin sheath and consists only of an axial cylinder and a Schwann sheath.
Function of nerve fibers.
The main function of nerve fibers is the transmission of nerve impulses. Currently, two types of nerve transmission have been studied: pulsed and non-pulse. Impulse transmission is provided by electrolyte and neurotransmitter mechanisms. The speed of nerve impulse transmission in myelinated fibers is much higher than in nonmyelinated fibers. In its implementation, the most important role is played by myelin. This substance is capable of isolating a nerve impulse, as a result of which signal transmission along the nerve fiber occurs spasmodically, from one node of Ranvier to another. Pulseless transmission is carried out by axoplasmic current along special axon microtubules containing trophogens - substances that have a trophic effect on the innervated organ.
Structural classification.
Based on the number and arrangement of dendrites and axons, neurons are divided into axonless neurons, unipolar neurons, pseudounipolar neurons, bipolar neurons, and multipolar (many dendritic arbors, usually efferent) neurons.
Axonless neurons are small cells grouped near the spinal cord in the intervertebral ganglia, which do not have anatomical signs of division of processes into dendrites and axons. All processes of the cell are very similar. The functional purpose of axonless neurons is poorly understood. Unipolar neurons are neurons with one process, present, for example, in the sensory nucleus of the trigeminal nerve in the midbrain.
Bipolar neurons are neurons that have one axon and one dendrite, located in specialized sensory organs - the retina, olfactory epithelium and bulb, auditory and vestibular ganglia;
Multipolar neurons - Neurons with one axon and several dendrites. This type of nerve cells predominates in the central nervous system. Pseudounipolar neurons are unique in their kind. One tip extends from the body, which immediately divides in a T-shape. This entire single tract is covered with a myelin sheath and is structurally an axon, although along one of the branches the excitation goes not from, but to the body of the neuron. Structurally, dendrites are branches at the end of this (peripheral) process. The trigger zone is the beginning of this branching (i.e., it is located outside the cell body).
Nerve fibers -
processes of nerve cells covered with membranes.
Some nerve fibers have a sheath made of a fat-like substance called myelin
. This substance performs trophic, protective and electrical insulating functions.
In the early stages of ontogenesis, the myelin sheath is absent; it develops in the first 2–3 years of life, its formation depends on the child’s living conditions. Under unfavorable conditions, the process of myelination can slow down for several years, which complicates the control and regulatory activity of the nervous system.
Nerve fibers combine with each other to form nerves
, which in the form of white threads are visible to the naked eye. Nerves connect all parts of our body with the central parts of the nervous system. The main function of nerve fibers and nerves is to conduct nerve impulses.
There are three types of nerves:
1. sensitive or centripetal - conduct nerve impulses to the central nervous system;
2. motor or centrifugal – conduct nerve impulses from the central nervous system to peripheral organs;
3. mixed - consist of sensory and motor fibers.
Glial cells (neuroglia)
more numerous than neurons, accounting for half the volume of the central nervous system. They are capable of division throughout their lives. Glial cells perform supporting, protective, insulating, and metabolic (supplying neurons with nutrients) functions.
During human development, the ratio between glial and nerve cells changes significantly. In a newborn, the number of neurons is higher than glial cells; by the age of 20–30, their ratio becomes equal; after 30 years, the number of glial cells increases.
Basic properties of nervous tissue.
1. Excitability
– the ability of nerve tissue cells to quickly respond to irritation by changing the electrical properties of the cell membrane and their metabolism.
A quantitative measure of excitability is the irritation threshold
– the minimum amount of stimulus that can cause a tissue response.
The most common and natural stimulus for all cells of the body is the nerve impulse
.
A stimulus of lesser strength is called subthreshold
, and a greater one is
suprathreshold.
The latter cause more significant response changes in the vital activity of the tissue or organism.
2. Conductivity –
the ability of living tissue to conduct excitation. Excitation occurs due to the spread of a nerve impulse, which passes through the synapse to neighboring cells and can be transmitted to any part of the nervous system.
Having arisen at the site of excitation, an action potential (a change in the electrical charge of the membrane) causes a change in electrical charges in the adjacent area, and those, in turn, in the next one, etc.
3. Lability –
the ability of excitable tissue to reproduce the maximum number of action potentials per unit time.
The functional state of nervous tissue depends on its lability. Pathological processes and fatigue lead to a decrease in lability, and systematic special training leads to its increase.
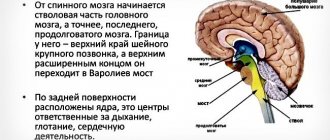
Anatomical and physiological features of the development of the central nervous system.
Spinal cord.
CM. of an adult is located in the spinal canal and is a cylindrical cord 40-45 cm long, with a total weight of 34 - 38 g. The spinal cord of a newborn is the most mature part of the central nervous system, but its final development ends only by the age of 20. During this period, the brain mass increases 8 times.
CM. has segmental
.
(31 segments). Two pairs of anterior and posterior roots extend from each segment. Each two pairs correspond to one vertebra. The dorsal roots are formed by sensory (afferent)
neurons.
Neurons located at the front of the spinal cord are motor neurons
, which control the functioning of skeletal muscles.
CM. conditionally divided into 4 departments:
1. cervical;
2. chest;
3. lumbar;
4. sacral,
Each of which contains several segments, a pair of spinal nerves extends from any segment. Each pair of nerves innervates a specific area of the body. For example: the nerves of the cervical and lumbar regions innervate the muscles of the limbs.
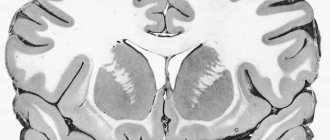
Building S.M. – cross section:
Gray matter located centrally
(nerve cells), which borders
the white matter
(nerve fibers). Sensitive roots s.m. consist of afferent neurons, and motor neurons - of efferent ones. Switching the signal from afferent to efferent neurons is carried out using intercalary or directly.
A huge number of reflex arcs are closed in the spinal cord, thanks to which it is able to regulate many functions of the body - such as flexion and extension of limbs, maintaining a certain posture...
Cm. human contains two thickenings: cervical and lumbar. They begin to develop in the first years of a child's life.
The cervical thickening regulates the movement of the upper limbs, the lumbar thickening regulates the movement of the lower limbs. The formation of cervical and lumbar thickenings depends on the child’s motor activity.
Nervous impulses from motor centers S.M. provides constant, slightly slower tension of the entire skeletal muscles, called muscle tone
, which allows a person to conduct normal motor activity.
Lloyd's classification[ | ]
Classifies only
afferent neurons.
| Fiber type | Function | Diameter, microns | Conduction speed, m/s | Myelination |
| Ia | Muscle spindles | 18-22 | 90-120 | + |
| Ib | Tendon receptors | 15-18 | 60-90 | — |
| II | Skin mechanoreceptors, secondary muscle spindles | 7-15 | 40-90 | + |
| III | Ligament receptors | 1-5 | 3-25 | intermittent |
| IV | Pain receptors, connective tissue receptors | 0,1-1 | 0,5-2 | — |
Myelination of nerve fibers[ | ]
During the formation of an unmyelinated nerve fiber, the axial cylinder (neuron process) is immersed in a cord of lemmocytes, the cytolemmas of which bend and tightly envelop the axial cylinder in the form of a coupling, the edges of which close above it, forming a duplication of the cell membrane - mesaxon
. Neighboring lemmocytes, which are part of a continuous glial cord, form simple contacts with their cytolemmas. Unmyelinated nerve fibers have weak insulation, allowing the transition of a nerve impulse from one fiber to another, both in the mesaxon area and in the area of interlemmocytic contacts.
Myelinated nerve fibers are much thicker than non-myelinated nerve fibers. The principle of formation of their membranes is the same as those without myelin, that is, the axial cylinders also bend the cytolemma of gliocytes, forming a linear mesaxon. However, the rapid growth of neurons in the somatic part of the nervous system, associated with the formation and growth of the entire organism, leads to the elongation of mesaxons and repeated circulation of lemmocytes around the axial cylinders. As a result, concentric layers are formed. In this case, the cytoplasm with the lemmocyte nucleus is pushed into the region of the last turn, forming the outer layer of the fiber sheath, called the Schwann sheath
or
neurilemma
.
The inner layer, consisting of mesaxon coils, is called myelin or myelin sheath. The consequence of the fact that myelination occurs during the growth of both neuron processes and lemmocytes themselves is a gradual increase in the number of turns and the size of the mesaxon, that is, each subsequent turn is wider than the previous one. Consequently, the last turn, containing the cytoplasm with the lemmocyte nucleus, is the widest. The thickness of myelin along the length of the fiber is heterogeneous, and at the points of contact of neighboring lemmocytes, the layered structure disappears and only the outer layers containing the cytoplasm and nucleus are in contact. The places of their contacts are called nodal nodes
(
intercepts of Ranvier
), which arise due to the absence of myelin here and thinning of the fiber.
In the central nervous system, myelination of the nerve fiber occurs due to the envelopment of the axial cylinders by the processes of oligodendrocytes.
As a membrane structure, myelin has a lipid base and, when treated with oxides, turns dark. Other components of the membrane and spaces are not stained, so light stripes are periodically encountered - myelin incisions
(
Schmidt-Lanterman notches
), which correspond to small layers of lemmocyte cytoplasm.
In the cytoplasm of the axial cylinder there are longitudinally oriented neurofibrils and mitochondria, which are more numerous in the immediate vicinity of the interceptions and in the terminal apparatus of the fiber. The cytolemma of the axial cylinder (axon) is called the axolemma
. It ensures the conduction of a nerve impulse, which is a wave of depolarization of the axolemma. If the axial cylinder is represented by a neurite, then it lacks granules of basophilic substance.
Neural mechanisms and effects on them. Part 2
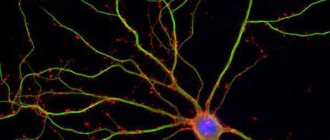
Individual nerve cells, or neurons, do not perform their functions as isolated units, like liver or kidney cells. The job of the 50 billion or so neurons in our brain is that they receive signals from some other nerve cells and transmit them to others.
Transmitting and receiving cells are organized into nerve circuits or networks. A single neuron with a divergent structure (from the Latin diverge - I deviate) can send signals to a thousand or even more other neurons. But more often, one such neuron connects to only a few specific neurons. In the same way, any neuron can receive input information from other neurons using one, several or many input connections if convergent paths converge on it (from the Latin converge - approaching, converging). Of course, everything depends on what kind of cell we are considering and what network it turned out to be included in during development. It is likely that only a small fraction of the pathways ending on a given neuron are active at any given time.
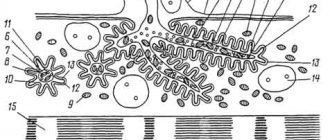
Rice. Nerve Network A large neuron with many dendrites receives information through synaptic contact with another neuron (in the upper left corner). The myelinated axon forms a synaptic contact with the third neuron (bottom). The surface of neurons is shown without the glial cells that surround the process directed towards the capillary (top right).
The complexity and diversity of the nervous system depends on the interactions between neurons, which in turn represent a set of different signals transmitted as part of the interaction of neurons with other neurons, muscles or glands. Signals are emitted and propagated by ions that generate an electrical charge that travels along the neuron.
The actual connection points - specific points on the surface of nerve cells where their contact occurs - are called synapses (synapsis; Greek “contact”, “connection”), and the process of transmitting information in these places is called synoptic transmission.
A synapse is a morphofunctional formation of the central nervous system, which ensures the transmission of a signal from a neuron to another neuron or to an effector cell (muscle, secretory). The synapse (point of contact) includes three components: the presynaptic membrane, the postsynaptic membrane and the synaptic cleft, i.e. contains elements of both the first and second contacting neurons. Some synapses cause depolarization of the neuron, others cause hyperpolarization; the former are excitatory, the latter are inhibitory. Typically, stimulation from several excitatory synapses is necessary to excite a neuron.
A synapse serves to transmit a nerve impulse between two cells, and during synaptic transmission the amplitude and frequency of the signal can be regulated. Some synapses cause depolarization of the neuron, others cause hyperpolarization; the former are excitatory, the latter are inhibitory. Typically, stimulation from several excitatory synapses is necessary to excite a neuron.
Most afferent inputs coming along the axons of other cells terminate at synapses on dendrites, but some terminate at synapses on the soma.
Excitatory nerve endings are often located distally on the dendrites, while inhibitory nerve endings are often located on the soma.
With some caveats, synapses can be compared to crossroads along the brain's pathways. At synapses, signals are transmitted in only one direction - from the terminal branch of the presynaptic neuron sending them to the nearest section of the postsynaptic neuron. However, the fast electrical transmission that works so well in the axon does not work in the synapse. Without going into the biological reasons for this, we can simply state that the chemical bond at synapses provides finer regulation of the properties of the postsynaptic cell membrane.
When communicating with each other, people convey the main content of their speech in words. To make more subtle accents or emphasize the additional meaning of words, they use the timbre of their voice, facial expressions, and gestures. During the communication of nerve cells, the main units of information are transmitted by specific chemical intermediaries - synaptic mediators (a certain neuron uses the same mediator in all its synapses). If we continue our analogy with verbal and nonverbal communication, we can say that some chemical intermediaries convey “facts,” while others convey additional semantic nuances or accents. Let's look further at mediators.
The process of a neuron - an axon - is directed to another neuron and forms a contact on it (axon-soma; axon-dendrite; axo-axonal and dendro-dendritic synapses are much less common). Breaking this contact, for example, with substances that block its operation, leads to serious violations. One of the main components are vesicles (vesicles), which contain a biologically very active substance called a neurotransmitter , or mediator (transmitter).
The main excitatory neurotransmitter in the brain is glutamate. For high accuracy of the nervous system, it is necessary to excite some neurons while suppressing the activity of others. This is achieved through braking. At inhibitory synapses, activation of receptors leads to the opening of ion channels allowing the release of anions, which leads to a change in membrane potential called inhibitory postsynaptic potential (IPSP). This prevents depolarization of the membrane, and hence the emergence of an action potential in the body of the receiving neuron. The main inhibitory neurotransmitters are gamma-aminobutyric acid (GABA) and glycine.
To find out the role of mediators, let's briefly look at the mechanism of synaptic transmission (this process will be discussed in more detail later).
Rice. Basic elements of a synapse
The nerve impulse travels along the fiber at high speed and approaches the synapse, which causes depolarization of the synapse membrane. However, this does not lead to the generation of new excitation (action potential), but causes the opening of special ion channels that allow calcium ions to pass into the synaptic cleft. In other words, the action potential arriving along the presynaptic fiber to the synapse causes depolarization of the membrane, which turns on the calcium pump, and calcium ions enter the synapse. Once in the cytoplasm of the synaptic ending (terminals), calcium enters into contact with proteins that form the shell of synaptic vesicles (vesicles), in which the transmitter is stored. The membranes of synaptic vesicles contract (the process of calcium binding by vesicle membrane proteins), pushing or injecting (exocytosis) their contents into the synaptic cleft between the membranes of two neurons. The excitation (electrical action potential) of a neuron at a synapse is converted from an electrical impulse to a chemical impulse. When a neuron is excited, a portion of a biologically active substance (transmitter) is released into the axon terminal and into the synaptic cleft. Next, the mediator molecules bind to special protein molecules - receptors located on the membrane of another neuron. Receptors match (bind) only one type of molecule (“one key fits one lock”).
The receptor consists of two parts: the “recognition” part (the mediator molecules have taken a position on the corresponding receptor molecule) and the “ion channel”, which opens for the entry of sodium ions or the exit of potassium ions from the cell. Thus, an ionic current flows through the membrane, which causes a change in the potential on the membrane (postsynaptic potential). In this case, the number of open channels is determined by the number of bound mediator molecules (and not by the potential on the membrane as in the case of an electrically excitable nerve fiber). The amplitude of the potential on the membrane is determined by the number of mediator molecules bound by the receptors.
Exocytosis (the cellular process by which a cell releases intracellular vesicles onto the outer cell membrane) can perform three main tasks:
- delivery to the cell membrane of a lipid necessary for cell growth;
- release of various compounds from the cell, such as toxic metabolic products or signaling molecules (hormones or neurotransmitters);
- delivery of functional membrane proteins to the cell membrane, such as receptors or transporter proteins. In this case, part of the protein that was directed inside the secreted vesicle turns out to be protruding outside the cell.
The following stages of endocytosis are distinguished (endocytosis is the process of capture (internalization) of external material by the cell, carried out through the formation of membrane vesicles, vesicles):
- Transportation of the vesicle from the place of synthesis and formation (Golgi apparatus - membrane structure) to the place of delivery is carried out by motor proteins along active filaments or microtubules of the cytoskeleton. This step may require movement of the secreted material over a considerable distance, as in a neuron. When the vesicle reaches the site of secretion, it comes into contact with specific retention factors of the cell membrane.
- Retention of the delivered vesicle is ensured by relatively weak bonds at a distance of more than 25 nm (nm is a nanometer; one nanometer is equal to 10–9 m; Here a meter is equal to the distance that light travels in a vacuum in a period of time equal to 1/299,792,458 of a second) and can serve , for example, to concentrate synaptic vesicles near the presynaptic membrane.
- Docking of the vesicle with the membrane is a direct continuation of the first phase of delivery, when the vesicle membrane comes into close contact with the cell membrane (5-10 nm). This involves the tight connection of the protein components of both membranes, caused by intramolecular rearrangements, and precedes the formation of the SNARE complex. (SNARE, English soluble NSF attachment receptor - a group of proteins that carry out the fusion of intracellular transport vesicles with the cell membrane (exocytosis) or target organelle).
- Stimulation (priming) of the vesicle actually corresponds to the formation of a special SNARE complex between two membranes and occurs only in the case of neuronal exocytosis. This stage includes processes of molecular rearrangements and ATP-dependent modifications of proteins and lipids (adenosine triphosphate, English ATP - a universal source of energy for all biochemical processes), occurring immediately before membrane fusion in response to an increase in the level of free calcium. This calcium-dependent process is necessary for rapid, controlled neurotransmitter release and is absent in the case of constitutive exocytosis.
- Fusion of the vesicle membrane with the cell membrane results in the release, or release, of the secreted vesicle contents into the extracellular space and the association of the vesicle's lipid bilayer with the outer membrane. In the case of synaptic release, the fusion process, as well as stimulation, is carried out by the SNARE complex.
Rice. Exocytosis at the synapse: signal transmission from neuron A to neuron B 1. Mitochondria; 2. Synaptic vesicle with a neurotransmitter; 3. Autoreceptor; 4. Synapse with a released neurotransmitter; 5. Postsynaptic receptor activated by a neurotransmitter; 6. Calcium channel; 7. Vesicle exocytosis; 8. Neurotransmitter recycling.
Rice. Scheme of transmitter release and processes occurring at the receiving synapse. 1. The synaptic vesicle releases the transmitter. 2. The mediator interacts with the receptor. The channel opens. 3. Movement of Na+ and K+ ions. 4. The presynaptic terminal reabsorbs the transmitter. 5. The antagonist blocks the receptor.
Let us consider the processes underlying the various changes produced by mediators in the cells on which they act. These changes are caused by ionic mechanisms associated with the electrical and chemical regulation of membrane properties. The excitability of a neuron changes because the transmitter changes the flow of ions moving into the cell or out of the cell. In order for ions to pass through the membrane, there must be holes in it. These are not just holes, but special large tubular proteins (receptors) called “channels”. Some of these channels are specific for a particular ion – sodium, potassium or calcium, for example; others are not so selective. Some channels can be opened by electrical commands (such as membrane depolarization at an action potential); others open and close under the influence of chemical messengers. Each chemical messenger affects cells through chemically mediated changes in ionic permeability. Certain ions and molecules used by one or another mediator therefore become the chemical equivalent of the transmitted signal.
Channels are protein molecules that “stitch” the membrane (one part of the molecule is located in the cytoplasm, and the other in the extracellular environment).
A neuron is capable of excitation, which consists in the fact that the neuron membrane in a resting state has a potential of the order of –70 mV (negativity in the cytoplasm), and in a state of excitation it acquires a potential of +55 mV. The absolute value of the action potential is about 125 mV. The duration of a neuron's action potential is about 1 ms (1/1000 s). Next, this excitation (action potential) must be transferred to another neuron or another cell (muscle, glandular, etc.).
Excitation in the form of an action potential leaves the body of the neuron along its process, which is called the axon. The axons of individual neurons have special covers made of myelin, a good electrical insulator, consisting of about two-thirds fat. Myelinated fibers conduct stimulation hundreds of times faster. Almost all nerve fibers in the human central nervous system have myelin sheaths and do not allow current to pass through their sections until they reach the nodes of Ranvier (at the junction of two myelin sheaths where there is no sheath). Consequently, excitation moves in leaps and bounds from interception to interception.
Action potential propagation along unmyelinated fibers. The AP propagates continuously along the unmyelinated fiber. The conduction of a nerve impulse begins with the propagation of an electric field. The resulting AP due to the electric field is capable of depolarizing the membrane of the neighboring area to a critical level, as a result of which new APs are generated in the neighboring area. The PDs themselves do not move; they disappear in the same place where they appear. The main role in the emergence of a new PD is played by the previous one.
If an intracellular electrode is used to stimulate the axon in the middle, the AP will propagate in both directions. Typically, the AP propagates along the axon in one direction (from the body of the neuron to the nerve endings), although depolarization of the membrane occurs on both sides of the area where the AP currently occurs. Unilateral conduction of AP is ensured by the properties of sodium channels - after opening, they are inactivated for some time and cannot open at any value of the membrane potential (refractory property). Therefore, in the area closest to the cell body, where the AP has already “passed” before, it does not occur.
All other things being equal, the propagation of APs along the axon occurs the faster, the larger the fiber diameter. The AP can propagate along the giant axons of the squid at almost the same speed as along the myelinated fibers of vertebrates (about 100 m/s).
Action potential propagation along myelinated fibers. Along the myelinated fiber, the AP propagates in a spasmodic manner (saltatory conduction). Myelinated fibers are characterized by a concentration of voltage-gated ion channels only in the areas of nodes of Ranvier; here their density is 100 times greater than in the membranes of unmyelinated fibers. There are almost no voltage-gated channels in the area of myelin couplings. An AP that arises in one node of Ranvier, due to the electric field, depolarizes the membrane of neighboring nodes to a critical level, which leads to the emergence of new APs in them, that is, the excitation passes spasmodically, from one node to another. If one node of Ranvier is damaged, the PD excites the 2nd, 3rd, 4th and even the 5th, since the electrical insulation created by the myelin couplings reduces the dissipation of the electric field. This increases the speed of AP propagation along myelinated fibers compared to unmyelinated fibers. In addition, myelinated fibers are thicker, and the electrical resistance of thicker fibers is less, which also increases the speed of impulse transmission along the myelinated fibers. Another advantage of saltatory conduction is its energy efficiency, since only nodes of Ranvier are excited, the area of which is less than 1% of the membrane, and, therefore, significantly less energy is needed to restore transmembrane gradients of Na+ and K+, consumed as a result of the occurrence of AP, which can have value at a high frequency of discharges traveling along the nerve fiber.
To imagine how effectively the conduction speed can be increased due to the myelin sheath, it is enough to compare the speed of impulse propagation through the unmyelinated and myelinated areas of the human nervous system. With a fiber diameter of about 2 µm and the absence of a myelin sheath, the conduction velocity will be ~1 m/s, and in the presence of even weak myelination with the same fiber diameter – 15-20 m/s. In fibers of larger diameter, which have a thick myelin sheath, the conduction speed can reach 120 m/s.
The speed of propagation of an action potential along the membrane of a single nerve fiber is by no means a constant value - depending on various conditions, this speed can decrease very significantly and, accordingly, increase, returning to a certain initial level.
The mediator acts for a very short time, after which it is destroyed by a specific enzyme. For example, in cholinergic synapses, the enzyme that destroys the transmitter in the synaptic cleft is acetylcholinesterase. At the same time, part of the transmitter can move through the postsynaptic membrane (direct uptake) and in the opposite direction through the presynaptic membrane (reverse uptake). In some cases, the mediator is also absorbed by neighboring neuroglial cells.
Structure and types of synapse Based on localization, synapses are divided into central and peripheral. The central ones are divided into axo-axonal, axo-dendritic, axo-somatic, dendro-dendritic, etc.; According to their development in ontogenesis, stable and dynamic synapses are distinguished; according to the final effect, inhibitory and excitatory synapses are distinguished. According to the mechanism of signal transmission, synapses can be electrical, chemical or mixed.
A typical synapse is axo-dendritic chemical. Such a synapse consists of two parts: presynaptic, formed by the club-shaped extension of the axon terminal of the transmitting cell, and postsynaptic, represented by the contacting area of the cytolemma of the receiving cell (in this case, the area of the dendrite). A synapse is a space separating the membranes of contacting cells to which nerve endings approach. The transmission of impulses is carried out chemically with the help of mediators or electrically through the passage of ions from one cell to another.
Between both parts there is a synaptic cleft - a gap 10–50 nm wide between the postsynaptic and presynaptic membranes, the edges of which are strengthened by intercellular contacts.
The part of the clavate extension axolemma adjacent to the synaptic cleft is called the presynaptic membrane. The section of the cytolemma of the receiving cell that borders the synaptic cleft on the opposite side is called the postsynaptic membrane; in chemical synapses it is prominent and contains numerous receptors.
In synaptic expansion there are small vesicles, so-called synaptic vesicles, containing either a mediator (a substance that mediates the transmission of excitation) or an enzyme that destroys this mediator. On the postsynaptic, and often on the presynaptic membranes, there are receptors for one or another mediator. Classifications of synapses
Depending on the mechanism of nerve impulse transmission, there are:
- chemical;
- electrical - cells are connected by highly permeable contacts using special connexons (each connexon consists of six protein subunits). The distance between cell membranes in the electrical synapse is 3.5 nm (usual intercellular distance is 20 nm)
Since the resistance of the extracellular fluid is low (in this case), impulses pass through the synapse without delay. Electrical synapses are usually excitatory.
Electrical synapses are less common in the mammalian nervous system than chemical ones.
- mixed synapses: The presynaptic action potential produces a current that depolarizes the postsynaptic membrane of a typical chemical synapse where the pre- and postsynaptic membranes are not tightly adjacent to each other. Thus, at these synapses, chemical transmission serves as a necessary reinforcing mechanism.
An electrical synapse is a gap-like formation with ion bridges-channels between two contacting cells. In the presence of an AP, the current almost unhindered jumps through the gap-like junction and induces the generation of an AP in another cell, thus rapid transfer of excitation occurs. But electrical synapses are mostly bidirectional. In addition, they cannot be used to force the effector cell to inhibit its activity. On the other hand, signal transmission occurs with almost no synaptic delay and almost no current leakage through the extracellular medium. Electrical synapses are widespread in the nervous systems of invertebrates and lower vertebrates. In the mammalian brainstem they are present in the trigeminal nerve nuclei and some other brainstem nuclei.
The most common are chemical synapses. Chemical synapses transmit nerve signals using chemicals - mediators, which are contained in synaptic vesicles.
Chemical synapses can be classified according to their location and belonging to the corresponding structures:
- peripheral
- neuromuscular
- neurosecretory (axo-vasal)
- receptor-neuronal
- central
- axo-dendritic - with dendrites, incl.
- axo-spinous - with dendritic spines, outgrowths on dendrites;
Chemical synapses are classified according to the nature of the transmitter they use:
- including adrenergic containing adrenaline or norepinephrine;
At the same time, only one transmitter is not always produced at the synapse. Usually the main pick is released along with another one that plays the role of a modulator.
According to the sign of action, synapses can be excitatory or inhibitory.
In the first case, one cell orders another to switch to activity, and in the second, on the contrary, it makes it difficult to activate the cell to which the signal is transmitted. Under constant inhibitory commands, some nerve cells remain silent until excitatory signals cause them to fire. For example, the nerve cells in the spinal cord that tell your muscles to act when you walk or dance are usually silent until they receive excitatory impulses from cells in the motor cortex. Under the influence of spontaneous excitatory commands, other nerve cells become active without waiting for conscious signals; for example, neurons that control the movements of the chest and diaphragm during breathing obey higher-level cells that respond only to the concentration of O2 and CO2 in the blood.
If the former contribute to the occurrence of excitation in the postsynaptic cell (in them, as a result of the arrival of an impulse, depolarization of the membrane occurs, which can cause an action potential under certain conditions), then the latter, on the contrary, stop or prevent its occurrence and prevent further propagation of the impulse. Typically inhibitory are glycinergic (mediator - glycine) and GABAergic synapses (mediator - gamma-aminobutyric acid).
Inhibitory synapses are of two types: 1) a synapse, in the presynaptic endings of which a transmitter is released, hyperpolarizing the postsynaptic membrane and causing the appearance of an inhibitory postsynaptic potential; 2) axo-axonal synapse, providing presynaptic inhibition. Cholinergic synapse (s. cholinergica) is a synapse in which acetylcholine is the mediator.
Some synapses have a postsynaptic seal, an electron-dense zone made of proteins. Based on its presence or absence, synapses are distinguished as asymmetric and symmetric. It is known that all glutamatergic synapses are asymmetrical, and GABAergic synapses are symmetrical.
In cases where several synaptic extensions come into contact with the postsynaptic membrane, multiple synapses are formed.
Special forms of synapses include spiny apparatus, in which short single or multiple protrusions of the postsynaptic membrane of the dendrite contact the synaptic extension. Spine apparatuses significantly increase the number of synaptic contacts on a neuron and, consequently, the amount of information processed. Non-spine synapses are called sessile synapses. For example, all GABAergic synapses are sessile.
Based on what science knows today, the interneuronal interactions that occur in the brain can largely be explained in terms of excitatory and inhibitory synaptic inputs. However, there are also more complex modifying influences that are of great importance, since they increase or decrease the intensity of the neuron's response to input signals from various other neurons.
Continued: Neural mechanisms and effects on them. Part 3