Для термина «волокно» см. также другие значения.
Не́рвные воло́кна
— длинные отростки нейронов, покрытые глиальными оболочками. По нервным волокнам распространяются нервные импульсы, по каждому волокну изолированно, не заходя на другие[1][2].
В различных отделах нервной системы оболочки нервных волокон значительно различаются по своему строению, что лежит в основе деления всех волокон на миелиновые
и
безмиелиновые
. Те и другие состоят из отростка нервной клетки, лежащего в центре волокна, и поэтому называемого
осевым цилиндром
(аксоном), и, в случае миелиновых волокон, окружающей его оболочкой. В зависимости от интенсивности функциональной нагрузки нейроны формируют тот или иной тип волокна. Для соматического отдела нервной системы, иннервирующей скелетную мускулатуру, обладающую высокой степенью функциональной нагрузки, характерен миелиновый (мякотный) тип нервных волокон, а для вегетативного отдела, иннервирующего внутренние органы — безмиелиновый (безмякотный) тип.
Покрытые оболочкой сплетения пучков нервных волокон образуют нервы.
Классификация Эрлангера-Гассера[ | ]
Является наиболее полной классификацией нервных волокон по скорости проведения нервного импульса.
| Тип волокна | Функция | Диаметр, мкм | Скорость проведения, м/с | Миелинизация |
| Aα | Афферентные — мышечные веретёна, сухожильные органы; эфферентные — скелетные мышцы | 10-20 | 60-120 | + |
| Aβ | Афферентные — тактильное чувство; коллатерали Aα волокон к интрафузальным мышечным волокнам | 7-15 | 40-90 | + |
| Aγ | Эфферентные — мышечные веретёна | 4-8 | 15-30 | + |
| Aδ | Афферентные — температура, быстрое проведение боли | 3-5 | 5-25 | + |
| B | Симпатические, преганглионарные; постганглионарные волокна цилиарного ганглия | 1-3 | 3-15 | прерывистая |
| C | Симпатические, постганглионарные; афферентные — медленное проведение боли | 0,3-1 | 0,5-2 | — |
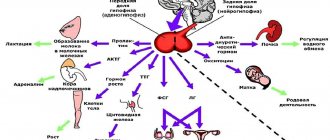
Строение и функции нервных волокон, их классификация
Нервные волокна — отростки нервных клеток (нейронов), имеющие оболочку и способные проводить нервный импульс.
Главной составной частью нервного волокна является отросток нейрона, образующий как бы ось волокна. Большей частью это аксон. Нервный отросток окружен оболочкой сложного строения, вместе с которой он и образует волокно.
Нервные волокна делятся на миелиновые и безмиелиновые. Первые имеют миелиновую оболочку, покрывающую аксон, вторые лишены миелиновой оболочки.
Как в периферической так и в центральной нервной системе преобладают миелиновые волокна. Нервные волокна, лишенные миелина располагаются преимущественно в симпатическом отделе вегетативной нервной системы.
Строение нервных волокон:
Миелиновое нервное волокно имеет в своём составе следующие элементы (структуры): 1) осевой цилиндр, располагающийся в самом центре нервного волокна,
2) миелиновую оболочку, покрывающую осевой цилиндр,
3) шванновскую оболочку.
Осевой цилиндр состоит из нейрофибрилл. Мякотная оболочка содержит большое количество веществ липоидного характера, известных под названием миелина. Миелин обеспечивает быстроту проведение нервных импульсов. Миелиновая оболочка покрывает осевой цилиндр не на всём промежутке, образуя промежутки, получившие название перехваты Ранвье. В области перехватов Ранвье осевой цилиндр нервного волокна примыкает к верхней — шванновской оболочке.
Промежуток волокна, расположенный между двумя перехватами Ранвье, называют сегментом волокна. В каждом таком сегменте на окрашенных препаратах можно видеть ядро шванновской оболочки. Оно лежит приблизительно посредине сегмента и окружено протоплазмой шванновской клетки, в петлях которой и содержится миелин. Между перехватами Ранвье миелиновая оболочка также не является сплошной. В толще ее обнаруживаются так называемые насечки Шмидт-Лантермана, идущие в косом направлении.
Клетки шванновской оболочки, так же как и нейроны с отростками, развиваются из эктодермы. Они покрывают осевой цилиндр нервного волокна периферической нервной системы аналогично тому, как клетки глии покрывают нервное волокно в центральной нервной системе. В результате этого они могут называться периферическими глиальными клетками.
В центральной нервной системе нервные волокна не имеют шванновских оболочек. Роль шванновских клеток здесь выполняют элементы олигодендроглии. Безмиелиновое (безмякотное) нервное волокно лишено миелиновой обкладки и состоит только из осевого цилиндра и шванновской оболочки.
Функция нервных волокон.
Главная функция нервных волокон – передача нервного импульса. В настоящее время изучено два типа нервной передачи: импульсная и безимпульсная. Импульсная передача обеспечивается электролитными и нейротрансмиттерными механизмами. Скорость передачи нервного импульса в миелиновых волокнах значительно выше, чем в безмякотных. В её осуществлении важнейшая роль принадлежит миелину. Данное вещество способно изолировать нервный импульс, в результате чего передача сигнала по нервному волокну происходит скачкообразно, от одного перехвата Ранвье к другому. Безимпульсная передача осуществляется током аксоплазмы по специальным микротрубочкам аксона, содержащим трофогены – вещества, оказывающие на иннервируемый орган трофическое влияние.
Структурная классификация.
На основании числа и расположения дейндритов и аксона нейроны делятся на безаксонные, униполярные нейроны, псевдоуниполярные нейроны, биполярные нейроны и мультиполярные (много дендритных стволов, обычно эфферентные) нейроны.
Безаксонные нейроны — небольшие клетки, сгруппированы вблизи спинного мозга в межпозвоночных ганглиях, не имеющие анатомических признаков разделения отростков на дендриты и аксоны. Все отростки у клетки очень похожи. Функциональное назначение безаксонных нейронов слабо изучено. Униполярные нейроны — нейроны с одним отростком, присутствуют, например в сенсорном ядре тройничного нерва в среднем мозге.
Биполярные нейроны — нейроны, имеющие один аксон и один дендрит, расположенные в специализированных сенсорных органах — сетчатке глаза, обонятельном эпителии и луковице, слуховом и вестибулярном ганглиях;
Мультиполярные нейроны — Нейроны с одним аксоном и несколькими дендритами. Данный вид нервных клеток преобладает в центральной нервной системе. Псевдоуниполярные нейроны — являются уникальными в своём роде. От тела отходит один остросток, который сразу же Т-образно делится. Весь этот единый тракт покрыт миелиновой оболочкой и структурно представляет собой аксон, хотя по одной из ветвей возбуждение идёт не от, а к телу нейрона. Структурно дендритами являются разветвления на конце этого (периферического) отростка. Триггерной зоной является начало этого разветвления (т. е. находится вне тела клетки).
Нервные волокна –
отростки нервных клеток, покрытые оболочками.
Некоторые нервные волокна имеют оболочку, состоящую из жироподобного вещества – миелина
. Это вещество выполняет трофическую, защитную и электроизолирующую функции.
На ранних этапах онтогенеза миелиновая оболочка отсутствует, она развивается в первые 2 – 3 года жизни, ее формирование зависит от условий жизни ребенка. В неблагоприятных условиях процесс миелинизации может замедляться на несколько лет, что затрудняет управляющую и регулирующую деятельность нервной системы.
Объединяясь друг с другом, нервные волокна образуют нервы
, которые в виде белых нитей видны невооруженным глазом. Нервы связывают все участки нашего тела с центральными отделами нервной системы. Основная функция нервных волокон и нервов – проведение нервных импульсов.
Различают три вида нервов:
1. чувствительные или центростремительные – проводят нервные импульсы в ЦНС;
2. двигательные или центробежные – проводят нервные импульсы от ЦНС к периферическим органам;
3. смешанные – состоят из чувствительных и двигательных волокон.
Глиальные клетки(нейроглии)
более многочисленны, чем нейроны, составляют половину объема ЦНС. Они способны к делению в течении всей жизни. Глиальные клетки выполняют опорную, защитную, изолирующую, обменную (снабжение нейронов питательными веществами) функции.
В процессе развития человека соотношение между глиальными и нервными клетками значительно меняется. У новорожденного количество нейронов выше, чем глиальных клеток, к 20 – 30 годам их соотношение становится равным, после 30 лет количество глиальных клеток увеличивается.
Основные свойства нервной ткани.
1. Возбудимость
– способность клеток нервной ткани быстро реагировать на раздражение посредством изменения электрических свойств мембраны клеток и их обмена веществ.
Количественной мерой возбудимости является порог раздражения
– минимальная величина раздражителя, способная вызвать ответную реакцию ткани. Наиболее общим и естественным раздражителем для всех клеток тела является
нервный импульс
. Раздражитель меньшей силы называют
подпороговым
, а большей –
надпороговым.
Последние вызывают более значительные ответные изменения в жизнедеятельности ткани или организма.
2. Проводимость –
способность живой ткани проводить возбуждение. Проведение возбуждения происходит за счет распространения нервного импульса, который переходит через синапс на соседние клетки и может передаваться в любой отдел нервной системы.
Возникши в месте возбуждения потенциал действия (изменение электрического заряда мембраны) вызывает изменение электрических зарядов в соседнем участке, а те в свою очередь в следующем и т.д.
3. Лабильность –
способность возбудимой ткани воспроизводить максимальное количество потенциалов действия в единицу времени.
Функциональное состояние нервной ткани зависит от ее лабильности. Патологические процессы и утомление приводят к снижению лабильности, а систематические специальные тренировки – к ее повышению.
Анатомо-физиологические особенности развития ЦНС.
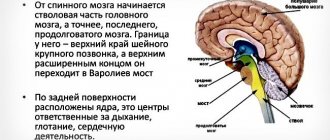
Спинной мозг.
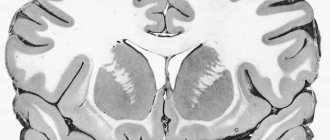
С.М. взрослого человека размещается в позвоночном канале и представляет собой цилиндрический тяж длиной 40-45 см, общей массой 34 – 38 г. Спинной мозг новорожденного является наиболее зрелой частью ЦНС, однако его окончательное развитие заканчивается только к 20 годам. За этот период масса мозга увеличивается в 8 раз.
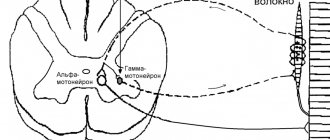
С.М. имеет сегментарное
. (31 сегмент). От каждого сегмента отходят по две пары передних и задних корешков. Каждые две пары соответствуют одному позвонку. Задние корешки образованы
чувствительными (афферентными)
нейронами. Нейроны, расположенные в передней части спинного мозга, являются
двигательными
, они управляют работой скелетных мышц.
С.М. условно разделяют на 4 отдела:
1. шейный;
2. грудной;
3. поясничный;
4. крестцовый,
Каждый из которых содержит несколько сегментов от любого сегмента отходит пара спинномозговых нервов. Каждая пара нервов иннервирует определенный участок организма. Например: нервы шейного и поясничного отделов иннервируют мышцы конечностей.
Строение С.М. – поперечный разрез:
Центрально расположено серое вещество
(нервные клетки), которое окаймляет
белое вещество
(нервные волокна). Чувствительные корешки с.м. состоят из афферентных нейронов, а двигательные – из эфферентных. Переключение сигнала с афферентных на эфферентные нейроны осуществляется с помощью вставочных или непосредственно.
В спинном мозге замыкается огромное количество рефлекторных дуг, благодаря этому он способен регулировать многие функции организма – такие как сгибание и разгибание конечностей, поддержание определенной позы….
С.м. человека содержит два утолщения: шейное и поясничное. Они начинают развиваться в первые годы жизни ребенка.
Шейное утолщение регулирует движение верхних конечностей, поясничное – нижних. Формирование шейного и поясничного утолщений зависит от двигательной активности ребенка.
Нервная импульсация из двигательных центров С.М. обеспечивает постоянное, чуть замедленное, напряжение всей скелетной мускулатуры, называемое мышечным тонусом
, что позволяет человеку вести нормальную двигательную деятельность.
Классификация по Ллойду[ | ]
Классифицирует только
афферентные нейроны.
| Тип волокна | Функция | Диаметр, мкм | Скорость проведения, м/с | Миелинизация |
| Ia | Мышечные веретёна | 18-22 | 90-120 | + |
| Ib | Сухожильные рецепторы | 15-18 | 60-90 | — |
| II | Механорецепторы кожи, вторичные мышечные веретёна | 7-15 | 40-90 | + |
| III | Рецепторы связок | 1-5 | 3-25 | прерывистая |
| IV | Болевые рецепторы, рецепторы соединительной ткани | 0,1-1 | 0,5-2 | — |
Миелинизация нервных волокон[ | ]
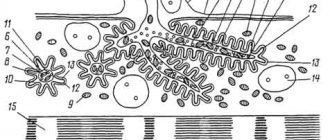
При формировании безмиелинового нервного волокна осевой цилиндр (отросток нейрона) погружается в тяж из леммоцитов, цитолеммы которых прогибаются и плотно охватывают осевой цилиндр в виде муфты, края которой смыкаются над ним, образуя дупликатуру клеточной мембраны — мезаксон
. Соседние леммоциты входящие в состав сплошного глиального тяжа своими цитолеммами образуют простые контакты. Безмиелиновые нервные волокна имеют слабую изоляцию, допускающую переход нервного импульса с одного волокна на другое, как в области мезаксона, так и в области межлеммоцитарных контактов.
Миелиновые нервные волокна значительно толще безмиелиновых. Принцип образования их оболочек такой же, как и безмиелиновых, то есть осевые цилиндры также прогибают цитолемму глиоцитов, образуя линейный мезаксон. Однако, быстрый рост нейронов соматического отдела нервной системы, связанный с формированием и ростом всего организма, приводит к вытягиванию мезаксонов, многократному обращению леммоцитов вокруг осевых цилиндров. В результате образуются концентрические наслоения. При этом цитоплазма с ядром леммоцитов оттесняется в область последнего витка, образующего наружный слой оболочек волокна, называемой шванновской оболочкой
или
неврилеммой
. Внутренний слой, состоящий из витков мезаксона, называется миелиновым или миелиновой оболочкой. Следствием того, что миелинизация происходит в процессе роста как отростков нейронов, так и самих леммоцитов, является постепенное увеличение количества витков и размеров мезаксона, то есть каждый последующий виток шире предыдущего. Следовательно, последний виток, содержащий цитоплазму с ядром леммоцита является самым широким. Толщина миелина по длине волокна неоднородна, а в местах контактов соседних леммоцитов слоистая структура исчезает и контактируют лишь наружные слои, содержащие цитоплазму и ядро. Места их контактов называются
узловыми перехватами
(
перехватами Ранвье
), возникающими вследствие отсутствия здесь миелина и истончения волокна.
В ЦНС миелинизация нервного волокна происходит за счет обхвата осевых цилиндров отростками олигодендроцитов.
Как мембранная структура миелин имеет липидную основу и при обработке окисями окрашивается в тёмный цвет. Другие компоненты мембраны и промежутки не окрашиваются, поэтому периодически встречаются светлые полоски − насечки миелина
(
насечки Шмидта-Лантермана
), которые соответствуют небольшим прослойкам цитоплазмы леммоцита.
В цитоплазме осевого цилиндра располагаются продольно ориентированные нейрофибриллы и митохондрии, которых больше в непосредственной близости к перехватам и в концевых аппаратах волокна. Цитолемма осевого цилиндра (аксона) называется аксолеммой
. Она обеспечивает проведение нервного импульса, который представляет собой волну деполяризации аксолеммы. Если осевой цилиндр представлен нейритом, то в нём отсутствуют гранулы базофильного вещества.
Нейронные механизмы и воздействие на них. Часть 2
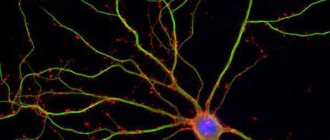
Отдельные нервные клетки, или нейроны, выполняют свои функции не как изолированные единицы, подобно клеткам печени или почек. Работа 50 миллиардов (или около того) нейронов нашего мозга состоит в том, что они получают сигналы от каких-то других нервных клеток и передают их третьим.
Передающие и принимающие клетки объединены в нервные цепи или сети. Отдельный нейрон с дивергентной структурой (от лат. diverge – отклоняюсь) может посылать сигналы тысяче и даже большему числу других нейронов. Но чаще один такой нейрон соединяется всего лишь с несколькими определенными нейронами. Точно так же какой-либо нейрон может получать входную информацию от других нейронов с помощью одной, нескольких или многих входных связей, если на нем сходятся конвергентные пути (от лат. converge – приближаюсь, схожусь). Конечно, все зависит от того, какую именно клетку мы рассматриваем и в какую сеть она оказалась включенной в процессе развития. Вероятно, в каждый момент времени активна лишь небольшая часть путей, оканчивающихся на данном нейроне.
Рис. Нервная сеть Крупный нейрон с множеством дендритов получает информацию, через синаптический контакт с другим нейроном (в левом верхнем углу). С помощью миелинизированного аксона образуется синаптический контакт с третьим нейроном (внизу). Поверхность нейронов изображена без клеток глии, которые окружают отросток, направленный к капилляру (справа вверху).
Сложность и многообразие нервной системы зависит от взаимодействия между нейронами, которые, в свою очередь, представляют собой набор различных сигналов, передаваемых в рамках взаимодействия нейронов с другими нейронами, мышцами или железами. Сигналы испускаются и распространяются с помощью ионов, генерирующих электрический заряд, который движется вдоль нейрона.
Действительные места соединения – специфические точки на поверхности нервных клеток, где происходит их контакт, – называются синапсами (synapsis; греч. «соприкосновение», «соединение»), а сам процесс передачи информации в этих местах – синоптической передачей.
Синапс – это морфофункциональное образование ЦНС, которое обеспечивает передачу сигнала с нейрона на другой нейрон или на эффекторную клетку (мышечную, секреторную). Синапс (место контакта) включает три компонента: пресинаптическая мембрана, постсинаптическая мембрана и синаптическая щель, т.е. содержит элементы и первого и второго контактирующих нейронов. Одни синапсы вызывают деполяризацию нейрона, другие – гиперполяризацию; первые являются возбуждающими, вторые – тормозящими. Обычно для возбуждения нейрона необходимо раздражение от нескольких возбуждающих синапсов.
Синапс служит для передачи нервного импульса между двумя клетками, причём в ходе синаптической передачи амплитуда и частота сигнала могут регулироваться. Одни синапсы вызывают деполяризацию нейрона, другие – гиперполяризацию; первые являются возбуждающими, вторые – тормозящими. Обычно для возбуждения нейрона необходимо раздражение от нескольких возбуждающих синапсов.
Большинство афферентных входов, поступающих по аксонам других клеток, оканчиваются синапсами на дендритах, но некоторые – синапсами на соме.
Возбуждающие нервные окончания чаще располагаются дистально на дендритах, а тормозные чаще находятся на соме.
С некоторыми оговорками синапсы можно сравнить с перекрестками на проводящих путях мозга. В синапсах сигналы передаются только в одном направлении – с концевой веточки посылающего их пресинаптического нейрона на ближайший участок постсинаптического нейрона. Однако быстрая электрическая передача, так хорошо действующая в аксоне, в синапсе не работает. Не вдаваясь в биологические причины этого, мы можем просто констатировать, что химическая связь в синапсах обеспечивает более тонкую регуляцию свойств мембраны постсинаптической клетки.
При общении друг с другом люди передают основное содержание своей речи словами. Чтобы сделать более тонкие акценты или подчеркнуть дополнительный смысл слов, они пользуются тембром голоса, мимикой, жестами. При коммуникации нервных клеток основные единицы информации передаются специфическими химическими посредниками – синоптическими медиаторами (определенный нейрон использует один и тот же медиатор во всех своих синапсах). Если продолжить нашу аналогию с вербальным и невербальным способом общения, то можно сказать, что одни химические посредники передают «факты», а другие – дополнительные смысловые оттенки или акценты. Рассмотрим далее медиаторы.
Отросток нейрона – аксон – направляется к другому нейрону и образует на нем контакт (аксон-сома; аксон-дендрит; значительно реже встречаются аксо-аксональные и дендро-дендритические синапсы.). Разрыв этого контакта, например, веществами, блокирующими его работу, приводит к серьезным нарушениям. Одним из основных компонентов являются пузырьки (везикулы), которые содержат биологически очень активное вещество, которое называется нейротрансмиттером, или медиатором (передатчиком).
Главный возбуждающий нейротрансмиттер в мозге – это глутамат. Для высокой точности работы нервной системы необходимо возбуждение одних нейронов при подавлении активности других. Это обеспечивается за счет торможения. В тормозных синапсах активация рецепторов ведет к открытию ионных каналов, обеспечивающих выход анионов, что приводит к изменению мембранного потенциала, называемому тормозным постсинаптическим потенциалом (ТПСП). Это препятствует деполяризации мембраны, а значит и возникновению потенциала действия в теле воспринимающего нейрона. Основными тормозными нейротрансмиттерами являются гамма-аминомасляная кислота (ГАМК) и глицин.
Чтобы выяснить роль медиаторов, бегло рассмотрим механизм синаптической передачи (далее этот процесс будет рассмотрен более детально).
Рис. Основные элементы синапса
Нервный импульс с большой скоростью продвигается по волокну и подходит к синапсу, что вызывает деполяризацию мембраны синапса. Однако это не приводит к генерации нового возбуждения (потенциала действия), а вызывает открывание специальных ионных каналов, которые пропускают ионы кальция в синаптическую щель. Иными словами, потенциал действия, пришедший по пресинаптическому волокну к синапсу, вызывает деполяризацию мембраны, которая включает кальциевый насос, и ионы кальция поступают в синапс. Попадая в цитоплазму синаптического окончания (терминали), кальций входит в связь с белками, образующими оболочку синапттических пузырьков (везикул), в которых хранится медиатор. Мембраны синаптических пузырьков сжимаются (процесс связывания кальция белками оболочки пузырька), выталкивая или впрыскивая (экзоцитоз) свое содержимое в синаптическую щель между мембранами двух нейронов. Возбуждение (электрический потенциал действия) нейрона в синапсе превращается из электрического импульса в химический импульс. При возбуждении нейрона порция биологически активного вещества (медиатора) выбрасывается в окончание аксона и в синаптическую щель. Далее молекулы медиатора связываются со специальными белковыми молекулами – рецепторами, находящимися на мембране другого нейрона. Рецепторы соответствуют (связывают) только один тип молекул («один ключ подходит к одному замку»).
Рецептор состоит из двух частей: «узнающая» часть (молекулы медиатора заняли позицию на соответствующей молекуле рецептора) и «ионный канал», который открывается для входа ионов натрия или выхода ионов калия из клетки. Таким образом, через мембрану протекает ионный ток, который вызывает изменение потенциала на мембране (постсинаптический потенциал). При этом количество открытых каналов определяется количеством связанных молекул медиатора (а не потенциалом на мембране как в случае электровозбудимого нервного волокна). Амплитуда потенциала на мембране определяется количеством молекул медиатора, связанного рецепторами.
Экзоцитоз (клеточный процесс, с помощью которого клетка выделяет внутриклеточные везикулы на внешнюю клеточную мембрану) может выполнять три основные задачи:
- доставка на клеточную мембрану липида, необходимого для роста клетки;
- высвобождение различных соединений из клетки, например токсичных продуктов метаболизма или сигнальных молекул (гормонов или нейромедиаторов);
- доставка функциональных мембранных белков на клеточную мембрану, таких как рецепторы или белки-транспортёры. При этом часть белка, которая была направлена внутрь секретируемой везикулы, оказывается выступающей снаружи клетки.
Различают следующие этапы эндоцитоза (endocytosis – процесс захвата (интернализации) внешнего материала клеткой, осуществляемый путём образования мембранных везикул, пузырьков):
- Транспортировка везикулы от места синтеза и формирования (аппарат Гольджи – мембранная структура) до места доставки осуществяется моторными белками вдоль активных филаментов либо микротрубочек цитоскелета. Этот этап может потребовать перемещения секретируемого материала на значительное расстояние, как, например, в нейроне. Когда везикула достигает места секреции, она входит в контакт со специфическими удерживающими факторами клеточной мембраны.
- Удержание доставленной везикулы обеспечивается относительно слабыми связями на расстоянии более 25 нм (нм – нанометр; один нанометр равен 10–9 м; Здесь метр равен расстоянию, которое проходит свет в вакууме за промежуток времени, равный 1/299 792 458 секунды) и может служить, например, для концентрирования синаптических везикул около пресинаптической мембраны.
- Стыковка везикулы с мембраной является непосредственным продолжением первой фазы доставки, когда мембрана везикулы входит в близкий контакт с мембраной клетки (5-10 нм). Это включает прочное соединение белковых компонентов обеих мембран, вызванное внутримолекулярными перестановками, и предваряет формирования SNARE комплекса. (SNARE, англ. soluble NSF attachment receptor – группа белков, осуществляющих слияние внутриклеточных транспортных везикул с клеточной мембраной (экзоцитоз) или органеллой-мишенью).
- Стимуляция (прайминг) везикулы фактически соответствует образованию особого SNARE-комплекса между двумя мембранами и осуществляется только в случае нейронального экзоцитоза. Этот этап включает процессы молекулярных перестановок и АТФ-зависимые модификации белков и липидов (аденозинтрифосфат, англ. АТР – универсальный источник энергии для всех биохимических процессов), происходящие непосредственно до слияния мембран в ответ на подъём уровня свободного кальция. Этот кальций-зависимый процесс необходим для быстрого контролируемого выброса нейромедиатора и отсутствует в случае конститутивного экзоцитоза.
- Слияние мембраны везикулы с мембраной клетки приводит к высвобождению, или выбросу, содержания секретируемой везикулы во внеклеточное пространство и объединению липидного бислоя везикулы с внешней мембраной. В случае синаптического выброса процесс слияния, так же как и стимуляция, осуществляется SNARE-комплексом.
Рис. Экзоцитоз в синапсе: передача сигнала от нейрона А к нейрону B 1. Митохондрия; 2. Синаптическая везикула с нейромедиатором; 3. Ауторецептор; 4. Синапс с выделенным нейромедиатором; 5. Постсинаптический рецептор, активируемый нейромедиатором; 6. Кальциевый канал; 7. Экзоцитоз везикулы; 8. Рециркуляция нейромедиатора.
Рис. Схема выброса медиатора и процессов, происходящих в принимающем синапсе. 1.Синаптический пузырек высвобождает медиатор. 2. Медиатор взаимодействует с рецептором. Канал открывается. 3. Перемещение ионов Na+ и К+. 4. Пресинаптическое окончание поглощает медиатор обратно. 5. Антагонист блокирует рецептор.
Рассмотрим процессы, лежащие в основе различных изменений, производимых медиаторами в клетках, на которые они воздействуют. Эти изменения обусловлены ионными механизмами, связанными с электрической и химической регуляцией свойств мембраны. Возбудимость нейрона изменяется потому, что медиатор изменяет поток ионов, переходящих внутрь клетки или же из клетки наружу. Для того чтобы ионы могли проходить через мембрану, в ней должны быть отверстия. Это не просто дыры, а специальные крупные трубчатые белки (рецепторы), называемые «каналами». Некоторые из этих каналов специфичны для определенного иона – натрия, калия или кальция, например; другие не столь избирательны. Некоторые каналы могут открываться с помощью электрических команд (таких, как деполяризация мембраны при потенциале действия); другие открываются и закрываются под действием химических посредников. Каждый химический посредник оказывает на клетки воздействие с помощью химически обусловленных изменений в ионной проницаемости. Определенные ионы и молекулы, используемые тем или иным медиатором, становятся поэтому химическим эквивалентом передаваемого сигнала.
Каналы представляют собой белковые молекулы, «прошивающие» мембрану (одна часть молекулы находится в цитоплазме, а другая во внеклеточной среде).
Нейрон способен к возбуждению, которое состоит в том, что мембрана нейрона в состоянии покоя имеет потенциал порядка –70мВ (отрицательность в цитоплазме), а в состоянии возбуждения приобретает потенциал +55 мВ. Абсолютная величина потенциала действия около 125 мВ. Длительность потенциала действия нейрона составляет около 1 мс (1/1000 с). Далее это возбуждение (потенциал действия) должно передаться другому нейрону или другой клетке (мышечной, железистой и др.).
Возбуждение в виде потенциала действия покидает тело нейрона по его отростку, который называется аксоном. Аксоны отдельных нейронов имеет специальные чехлы из миелина, хорошего электрического изолятора, состоящего примерно на две трети из жира. Миелинизированные волокна проводят возбуждение в сотни раз быстрее. Практически все нервные волокна в центральной нервной системе человека имеют миелиновые чехлы и не пропускают ток на своих участках до выхода на перехваты Ранвье (на стыке двух участков миелина, где нет покрытия). Следовательно, возбуждение движется скачками от перехвата к перехвату.
Распространение потенциала действия по немиелинизированным волокнам. По немиелинизированному волокну ПД распространяется непрерывно. Проведение нервного импульса начинается с распространением электрического поля. Возникший ПД за счет электрического поля способен деполяризовать мембрану соседнего участка до критического уровня, в результате чего на соседнем участке генерируются новые ПД. Сами ПД не перемещаются, они исчезают там же, где возникают. Главную роль в возникновении нового ПД играет предыдущий.
Если внутриклеточным электродом раздражать аксон посередине, то ПД будет распространяться в обоих направлениях. Обычно же ПД распространяется по аксону в одном направлении (от тела нейрона к нервным окончаниям), хотя деполяризация мембраны происходит по обе стороны от участка, где в данный момент возник ПД. Одностороннее проведение ПД обеспечивается свойствами натриевых каналов – после открывания они на некоторое время инактивируются и не могут открыться ни при каких значениях мембранного потенциала (свойство рефрактерности). Поэтому на ближнем к телу клетки участке, где до этого уже «прошел» ПД, он не возникает.
При прочих равных условиях распространение ПД по аксону происходит тем быстрее, чем больше диаметр волокна. По гигантским аксонам кальмара ПД может распространяться почти с такой же скоростью, как и по миелинизированным волокнам позвоночных (около 100 м/c).
Распространение потенциала действия по миелинизированным волокнам. По миелинизированному волокну ПД распространяется скачкообразно (сальтаторное проведение). Для миелинизированных волокон характерна концентрация потенциалзависимых ионных каналов только в областях перехватов Ранвье; здесь их плотность в 100 раз больше, чем в мембранах безмиелиновых волокон. В области миелиновых муфт потенциалзависимых каналов почти нет. ПД, возникший в одном перехвате Ранвье, за счет электрического поля деполяризует мембрану соседних перехватов до критического уровня, что приводит к возникновению в них новых ПД, то есть возбуждение переходит скачкообразно, от одного перехвата к другому. В случае повреждения одного перехвата Ранвье ПД возбуждает 2-й, 3-ий, 4-й и даже 5-й, поскольку электроизоляция, создаваемая миелиновыми муфтами, уменьшает рассеивание электрического поля. Это увеличивает скорость распространения ПД по миелинизированным волокнам по сравнению с немиелинизированными. Кроме того, миелинизированные волокна толще, а электрическое сопротивление более толстых волокон меньше, что тоже увеличивает скорость проведения импульса по миелинизированным волокнам. Другим преимуществом сальтаторного проведения является его экономичность в энергетическом плане, так как возбуждаются только перехваты Ранвье, площадь которых меньше 1 % мембраны, и, следовательно, необходимо значительно меньше энергии для восстановления трансмембранных градиентов Na+ и K+, расходующихся в результате возникновения ПД, что может иметь значение при высокой частоте разрядов, идущих по нервному волокну.
Чтобы представить, насколько эффективно может быть увеличена скорость проведения за счёт миелиновой оболочки, достаточно сравнить скорость распространения импульса по немиелинизированным и миелинизированным участкам нервной системы человека. При диаметре волокна около 2 µм и отсутствии миелиновой оболочки скорость проведения будет составлять ~1 м/с, а при наличии даже слабой миелинизации при том же диаметре волокна – 15-20 м/с. В волокнах большего диаметра, обладающих толстой миелинововой оболочкой, скорость проведения может достигать 120 м/с.
Скорость распространения потенциала действия по мембране отдельно взятого нервного волокна отнюдь не является постоянной величиной – в зависимости от различных условий, эта скорость может очень значительно уменьшаться и, соответственно, увеличиваться, возвращаясь к некоему исходному уровню.
Медиатор действует в течение очень короткого времени, после чего разрушается специфическим ферментом. Например, в холинэргических синапсах фермент, разрушающий медиатор в синаптической щели – ацетилхолинэстераза. Одновременно часть медиатора может перемещаться через постсинаптическую мембрану (прямой захват) и в обратном направлении – через пресинаптическую мембрану (обратный захват). В ряде случаев медиатор также поглощается соседними клетками нейроглии.
Структура и виды синапса По локализации синапсы делятся на центральные и периферические. Центральные подразделяются на аксо-аксональные, аксо-дендрические, аксо-соматические, дендро-дендрические и т.д.; по развитию в онтогенезе различают стабильные и динамические синапсы, по конечному эффекту – тормозные и возбуждающие. По механизму передачи сигнала синапсы бывают электрические, химические и смешанные.
Типичный синапс – аксо-дендритический химический. Такой синапс состоит из двух частей: пресинаптической, образованной булавовидным расширением окончанием аксона передающей клетки и постсинаптической, представленной контактирующим участком цитолеммы воспринимающей клетки (в данном случае – участком дендрита). Синапс представляет собой пространство, разделяющее мембраны контактирующих клеток, к которым подходят нервные окончания. Передача импульсов осуществляется химическим путём с помощью медиаторов или электрическим путём посредством прохождения ионов из одной клетки в другую.
Между обеими частями имеется синаптическая щель – промежуток шириной 10–50 нм между постсинаптической и пресинаптической мембранами, края которой укреплены межклеточными контактами.
Часть аксолеммы булавовидного расширения, прилежащая к синаптической щели, называется пресинаптической мембраной. Участок цитолеммы воспринимающей клетки, ограничивающий синаптическую щель с противоположной стороны, называется постсинаптической мембраной, в химических синапсах она рельефна и содержит многочисленные рецепторы.
В синаптическом расширении имеются мелкие везикулы, так называемые синаптические пузырьки, содержащие либо медиатор (вещество-посредник в передаче возбуждения), либо фермент, разрушающий этот медиатор. На постсинаптической, а часто и на пресинаптической мембранах присутствуют рецепторы к тому или иному медиатору. Классификации синапсов
В зависимости от механизма передачи нервного импульса различают:
- химические;
- электрические – клетки соединяются высокопроницаемыми контактами с помощью особых коннексонов (каждый коннексон состоит из шести белковых субъединиц). Расстояние между мембранами клетки в электрическом синапсе – 3,5 нм (обычное межклеточное – 20 нм)
Так как сопротивление внеклеточной жидкости мало (в данном случае), импульсы проходят не задерживаясь через синапс. Электрические синапсы обычно бывают возбуждающими.
Для нервной системы млекопитающих электрические синапсы менее характерны, чем химические.
- смешанные синапсы: Пресинаптический потенциал действия создает ток, который деполяризует постсинаптическую мембрану типичного химического синапса, где пре- и постсинаптические мембраны не плотно прилегают друг к другу. Таким образом, в этих синапсах химическая передача служит необходимым усиливающим механизмом.
Электрический синапс представляет собой щелевидное образование с ионными мостиками-каналами между двумя контактирующими клетками. При наличии ПД ток почти беспрепятственно перескакивает через щелевидный контакт и индуцирует генерацию ПД в другой клетке, таким образом происходит быстрая передача возбуждения. Но электрические синапсы в основном обладают двусторонней проводимостью. Кроме того с их помощью нельзя заставить эффекторную клетку тормозить свою активность. С другой стороны, передача сигнала происходит почти без синаптической задержки и почти без утечки токов через внеклеточную среду. Электрические синапсы широко распространены в нервной системе беспозвоночных и низших позвоночных. В стволе мозга млекопитающих они имеются в ядрах тройничного нерва и некоторых других ядрах ствола.
Наиболее распространены химические синапсы. Химические синапсы передачу нервного сигнала осуществляют с помощью химических веществ – медиаторов, которые содержатся в синаптических визикулах.
Химические синапсы можно классифицировать по их местоположению и принадлежности соответствующим структурам:
- периферические
- нервно-мышечные
- нейросекреторные (аксо-вазальные)
- рецепторно-нейрональные
- центральные
- аксо-дендритические – с дендритами, в т. ч.
- аксо-шипиковые – с дендритными шипиками, выростами на дендритах;
Химические синапсы классифицируют по природе медиатора, который они используют:
- в том числе адренергические, содержащие адреналин или норадреналин;
При этом в синапсе не всегда вырабатывается только один медиатор. Обычно основной медиатор выбрасывается вместе с другим, играющим роль модулятора.
По знаку действия синапсы могут быть возбуждающие и тормозные.
В первом случае одна клетка приказывает другой переходить к активности, а во втором, наоборот, затрудняет активацию клетки, которой передается сигнал. Под действием постоянных тормозящих команд некоторые нервные клетки хранят молчание до тех пор, пока возбуждающие сигналы не заставят их активироваться. Например, нервные клетки спинного мозга, которые приказывают вашим мышцам действовать, когда вы ходите или танцуете, обычно «молчат», пока не получат возбуждающих импульсов от клеток моторной коры. Под действием спонтанных возбуждающих команд другие нервные клетки переходят к активности, не дожидаясь осознанных сигналов; например, нейроны, управляющие движениями грудной клетки и диафрагмы при дыхании, подчиняются клеткам более высокого уровня, которые реагируют только на концентрацию О2 и СО2 в крови.
Если первые способствуют возникновению возбуждения в постсинаптической клетке (в них в результате поступления импульса происходит деполяризация мембраны, которая может вызвать потенциал действия при определённых условиях.), то вторые, напротив, прекращают или предотвращают его появление, препятствуют дальнейшему распространению импульса. Обычно тормозными являются глицинергические (медиатор – глицин) и ГАМК-ергические синапсы (медиатор – гамма-аминомасляная кислота).
Тормозные синапсы бывают двух видов: 1) синапс, в пресинаптических окончаниях которого выделяется медиатор, гиперполяризующий постсинаптическую мембрану и вызывающий возникновение тормозного постсинаптического потенциала; 2) аксо-аксональный синапс, обеспечивающий пресинаптическое торможение. Синапс холинергический (s. cholinergica) – синапс, медиатором в котором является ацетилхолин.
В некоторых синапсах присутствует постсинаптическое уплотнение – электронно-плотная зона, состоящая из белков. По её наличию или отсутствию выделяют синапсы асимметричные и симметричные. Известно, что все глутаматергические синапсы асимметричны, а ГАМКергические – симметричны.
В случаях, когда с постсинаптической мембраной контактирует несколько синаптических расширений, образуются множественные синапсы.
К специальным формам синапсов относятся шипиковые аппараты, в которых с синаптическим расширением контактируют короткие одиночные или множественные выпячивания постсинаптической мембраны дендрита. Шипиковые аппараты значительно увеличивают количество синаптических контактов на нейроне и, следовательно, количество перерабатываемой информации. «Не-шипиковые» синапсы называются «сидячими». Например, сидячими являются все ГАМК-ергические синапсы.
Судя по тому, что сегодня известно науке, межнейронные взаимодействия, происходящие в мозгу, можно в основном объяснить, исходя из возбуждающих и тормозных синаптических воздействий. Однако существуют и более сложные модифицирующие воздействия, которые имеют большое значение, так как они усиливают или уменьшают интенсивность ответной реакции нейрона на входные сигналы от различных других нейронов.
Продолжение: Нейронные механизмы и воздействие на них. Часть 3