| Finite brain | |
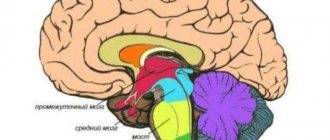
| Diagram depicting the major subdivisions of the embryonic vertebral brain. The forebrain in the five-vesicle stage is divided into the telencephalon and diencephalon | |
| Latin name | telencephalon |
| Catalogs |
|
| Telencephalon at Wikimedia Commons | |
Finite brain
(lat. telencephalon) - the most anterior part of the brain.
It consists of two cerebral hemispheres
(covered by the cortex), the corpus callosum, the striatum and the olfactory brain[1]. It is the largest section of the brain. It is also the most developed structure, covering all parts of the brain.
The cerebrum (lat. cerebrum) consists of two hemispheres, each of which is represented by the cortex, the olfactory brain and the basal ganglia. The cavity of the telencephalon is the lateral ventricles, located in each hemisphere. The hemispheres of the cerebrum are separated from each other by the longitudinal fissure of the cerebrum and are connected by the corpus callosum, the anterior and posterior commissures and the commissure of the fornix.
The corpus callosum consists of transverse fibers, which continue in the lateral direction into the hemispheres, forming the radiation of the corpus callosum, connecting parts of the frontal and occipital lobes of the hemispheres with each other, bending in an arcuate manner and forming the anterior - frontal and posterior - occipital forceps. Adjacent to the posterior and middle parts of the corpus callosum below is the fornix of the brain, consisting of two arched cords fused in its middle part with the help of the anterior commissure of the brain.
The importance of smell for mammals[ | ]
The olfactory organs of mammals are more developed than those of other terrestrial vertebrates, and play a very important role in their lives. Mammals use the ability to distinguish smells for orientation in space, when searching for food, and within the framework of interspecific and intraspecific contacts[4]. The importance of smell for mammals is also evidenced by the fact that the most extensive family of genes[en] in their genome is formed by just the genes that code olfactory receptor proteins[5]. The sense of smell also plays a certain role in the feeding of mammals: pleasant odors trigger the secretion of saliva and gastric juice, while unpleasant odors warn of potential harm (for example, the unpleasant smell of spoiled food) [6].
Based on the degree of development of the olfactory function, mammals are divided into two groups: macrosmates
with an exceptionally acute sense of smell (most mammals) and
microsmates
with a moderately developed sense of smell (primates, cetaceans, pinnipeds)[7].
The dog's nose is a hypersensitive organ of smell.
The difference between these groups is clearly visible when comparing the degree of development of the sense of smell in humans and dogs. If the human nose has about 6 million olfactory cells, then the dog has about 225 million [8]. Many macrosmats sense odors at a distance of several hundred meters and are able to find food objects underground[9]. The practice of searching for truffles growing underground in the forest with the help of specially trained search dogs and pigs, which are able to smell truffles underground at a distance of up to 20 m, is well known[10].
The degree of development of the sense of smell generally correlates with the number of genes that express various types of functional proteins of olfactory receptors. In macrosmates there are usually more than 1000 of them, in many primates - about 500, in humans - only 387 (about 1-2% of the genome [11]), in the platypus - 262 [12]. Apparently, the sense of smell is worst developed in cetaceans; They also have the highest percentage of olfactory receptor pseudogenes [13].
Functional anatomy of the telencephalon
The telencephalon consists of three parts that arose in different periods:
- Olfactory brain
- basal ganglia,
- Cloak (mantle).
Also inside the telencephalon are two lateral ventricles (the largest).
Olfactory brain: peripheral and central parts
The olfactory brain appeared with the development of the olfactory organs. Divided by:
- Peripheral department (below the central):
- Olfactory bulbs,
- Olfactory tracts
- Olfactory triangles
- Anterior perforated substance;
- Central section (on the medial surface of the hemispheres + deep in the lateral ventricles):
- vaulted gyrus,
- Hippocampus,
- Dentate gyrus.
What is included in the basal ganglia? Their localization
The basal ganglia is a collection of gray matter within the telencephalon, surrounded by white matter.
1) The striatum - consists of alternating areas of gray and white matter.
- caudate nucleus,
- Lenticular nucleus: shell (lateral part),
- globus pallidus (lateral and medial nuclei (lamellae)).
2) The fence is the shape of a thin plate (lateral to the lenticular nucleus).
3) Amygdala - basal nuclei, located in the temporal lobe of the hemispheres (closer to the lower surface).
Connections between banal kernels and functions
Conducting pathways of the basal ganglia:
- Internal connections - connect individual basal ganglia (some are permanent, others are formed individually).
- Afferent connections (coming from all parts of the brain).
- Efferent connections are outgoing connections to those parts of the GM.
Functions of the basal ganglia:
- Instincts are unconditioned reflexes;
- Complex automatic movements;
- Autonomic functions (basal ganglia are part of the limbic system);
- Feelings and emotions;
- Formation of the striopallidal system (coordinated work of the pyramidal and extrapyramidal systems: initially, movements are controlled by the pyramidal system => then, as skills are acquired, the extrapyramidal system).
The striopallidal system consists of:
- caudate nucleus,
- putamen (lateral part of the lenticular nucleus),
- fence.
- Formation of the pallidal system . It consists (also) of the midbrain formations:
- globus pallidus (part of the lentiform nucleus),
- substantia nigra (midbrain),
- red nuclei (midbrain).
Difference between the mantle and other structures of the telencephalon, 3 stages of development of the cortex
The cloak covers all other parts of the telencephalon. Consists of gray and white matter.
Gray matter - the cortex, is represented by clusters of neuron bodies lying on the surface (above the white matter).
White matter - pathways.
Three stages of cortex development:
- The ancient cortex - the cells above the white matter - are not organized and are located chaotically. Located on the basal surface, closer to the amygdala.
- Old cortex - the appearance of elements of ordered nerve cells by similarity and function. Layers are formed (no more than three). It is located on the medial surface, next to the central part of the olfactory brain.
- New cortex - cells are strictly ordered, differ in shape and function. Form layers (6 layers). Cells in different areas are different: Cytoarchitectonic fields (about 130),
- Myelarchitectonic fields (100) - differences in the structure of processes,
- Neuroarchitectonic fields (80) - along neuroglia,
- Angioarchitectonic fields - according to the vascular pattern.
Where is the new crust located and how does it differ from the ancient and old crust?
The neocortex covers 90% of the entire surface of the telencephalon. It is characterized by:
- Clear layered structure (6 layers),
- Vertical orientation, impulses travel through layers,
- Specialization of each layer according to functions,
- Presence of architectonic fields.
What functions are inherent in the cortex? What is analysis and synthesis?
- Analytical function (analysis) is the decomposition of objects and phenomena into individual characteristics;
- Synthetic function (synthesis) - from individual features - ideas about an object or phenomenon.
What 3 morphological substrates provide analytical and synthetic function?
Cloak functions:
1) Analytical - decomposition of objects and phenomena into individual characteristics.
For this purpose, there is a cortical center - a section of the cortex that contributes to the perception of one sign.
The bark itself consists of:
- Cortical nucleus (contains cells specialized in one trait);
- The scattered part (contains cells that perform the sensation of one sign, but can be rearranged to another sign).
2) Synthetic function - an idea of an object or phenomenon is made up of individual features. Synthetic functions perform:
- Associative fields are the zone of intersection of the peripheral (scattered) parts of several cortical centers. Here are cells that are initially capable of performing two functions.
- Association nerve fibers.
Where are the centers located: hearing, vision, taste, smell, skin sense, motor analyzer, balance?
Centers of the first signaling system (the system of the biological essence of man, the biological functions of which are found in other animals).
- The center of hearing is the superior temporal gyrus,
- The center of the cutaneous sense (tactile sense is the superior parietal lobule,
- Temperature and pain sensation – posterior postcentral gyrus,
- The center of stereognosis is the paracentral lobule,
- Proprioceptive sense – anterior central gyrus (3rd layer of cortex),
- Center of vision - along the banks of the calcarine groove (bird's spur groove),
- The center of taste and smell is the hook,
- The center of the motor analyzer is the anterior central gyrus (layer 5 of the cortex),
- The center of practical skills is in the area of the supramarginal gyrus.
Localization of centers of the second signaling system
The second signaling system is the provision of higher nervous activity in humans. All centers are reduced to human speech:
- The center of speech articulation is in the posterior part of the inferior frontal gyrus;
- The center of writing and written speech is in the posterior part of the middle frontal gyrus;
- The center for the perception of oral speech is the upper section of the superior temporal gyrus;
- The center of reading and perception of written speech is the angular gyrus.
The centers of the second signaling system are located next to those similar in function to the first signaling system.
Integrative function of the cortex: what is it based on, what is it ensured? Where do the descending tracts from the cortex go?
The cerebral cortex unites the entire nervous system into a single whole, and through the nervous system - the entire organism.
The integrative function is based on projection pathways. They connect the telencephalon with the underlying parts of the brain. This is accomplished through a complex reflex arc. Includes two entities:
- Internal capsule - includes all projection paths;
- radiant crown.
What is the limbic system?
- Central part of the olfactory brain
- amygdala complex,
- transparent partition,
- vault,
- Hippocampus,
- Orbital crust.
All this is the final brain.
Connections to the limbic system
- Internal (short) connections - pass through two circles of Papets (large, small circle)
Impulses through the internal connections of the limbic system flow into the hippocampus and amygdala complex - this is the core of the limbic system.
- External (long) connections:
- Double sided;
- Communication with all cortical centers;
- With all associative fields of the cortex (integrative areas of the cortex);
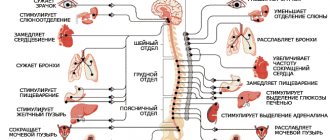
- Bilateral communication with the hypothalamus, reticular formation, brain stem.
In addition, the drug receives collaterals from all sensitive pathways.
Functions of the limbic system
- Instinctive behavior (in humans, instincts are controlled by the cortex),
- Autonomic function (visceral brain),
- Endocrine function,
- Sleep and wakefulness
- Memory (long-term and short-term),
- Attention (active and passive),
- emotional status,
- Determination of human gender (amygdala complex).
3 0
Source: https://medfsh.ru/teoriya/teoriya-po-anatomii/voprosy-po-anatomii/funktsionalnaya-anatomiya-konechnogo-mozga
Olfactory organs[ | ]
The olfactory system of mammals using the example of a human: 1 - olfactory bulb, 2 - mitral cells[en], 3 - bone, 4 - nasal epithelium, 5 - olfactory glomerulus[en], 6 - olfactory receptors The olfactory organs of
mammals are located in the posterior upper part of the nasal cavity cavities where a very complex system of olfactory nasal conchae arises, especially in macrosmates[en]
- thin bony lobes directed into the cavity and covered with olfactory epithelium. In the nasal concha, not only does the olfactory analysis of inhaled air take place, but also it is heated before entering the lungs. Among modern species of tetrapods, only mammals have olfactory shells, as well as a few species of birds in which these shells arose independently [14]. The olfactory epithelium contains olfactory receptor cells, supporting cells that secrete mucus and have properties similar to glial cells, as well as basal cells, which, like stem cells, are able to divide and give rise to new functional neurons throughout the life of the animal. The size of the olfactory epithelium in mammals varies from 2-4 cm² (human) and 9.3 cm² (rabbit) to 18 cm² (dog) and 21 cm² (domestic cat). However, these values do not give an idea of the acuity of smell, because they do not take into account the number of olfactory receptors per unit surface. Olfactory receptors are capable of detecting odorant molecules contained in the inhaled air. Like taste receptors, they belong to the group of chemoreceptors. Signals about the presence of odorous substances are transmitted through the olfactory nerve to the corresponding center of the brain - the olfactory bulb, or the primary olfactory centers of the cerebral cortex. From the latter, olfactory signals are transmitted to the hypothalamus, limbic system, reticular formation and neocortex [7][6][15].
Most mammals retain the Jacobson's organ as a separate section of the olfactory capsule. This organ, which is also present in bipeds and most tetrapods (the most important exceptions are birds and molehills), serves mainly for the perception of pheromones. In representatives of a number of groups (cetaceans, sirenians, most bats and narrow-nosed primates, including humans), the Jacobson organ is rudimentary or completely lost [16][17][18].
The vomeronasal organ is lined by an olfactory epithelium similar to that covering the turbinates. Olfactory receptor cells are renewed throughout life and are supported by epithelial and basal cells, but instead of cilia they have microvilli (microvilli). Receptor molecules are also represented by GPCRs, but their amino acid sequence has nothing in common with the turbinate receptors. Vomeronasal organ GPCRs are represented by two distinct families, each containing 100 to 200 genes and evolving independently. Members of one of these families have a long extracellular N-terminal domain similar to the metabotropic glutamate receptor. The secondary messenger here is not cAMP, as in the nasal turbinates, but inositol triphosphate. Afferent fibers from the vomeronasal epithelium project to the accessory olfactory bulb, which in most is located posterior to the main olfactory bulb. Like the olfactory epithelium of the nose, the vomeronasal epithelium is also divided into zones: different G proteins are expressed in the apical and basal parts of the organ. These zones are also preserved in projections into the additional olfactory bulb: the apical zone of the epithelium is projected into the anterior zone of the bulb, and the basal zone into the posterior zone. Glomeruli in the additional bulb are less pronounced than in the main one. In addition, instead of a spatial map of the main bulb, the representation of the additional bulb is more complex and mosaic. The additional bulb has no projections to the cortex and is connected only with the limbic system: with the amygdala and hypothalamic nuclei, which play an important role in sexual behavior. Perhaps the additional bulb reacts only to special species-specific combinations of substances of the corresponding pheromone and simply ignores all the others [19].
Functions
The name of the structure itself indicates what it is intended for. The functions of the olfactory nerve are to capture smell and decipher it. They cause appetite and salivation if the aroma is pleasant, or, on the contrary, they provoke nausea and vomiting when the amber leaves much to be desired.
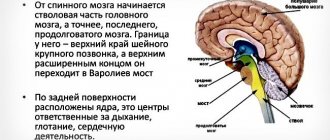
In order to achieve this effect, the olfactory nerve passes through the reticular formation and is directed to the brain stem. There the fibers connect to the nuclei of the intermediate, glossopharyngeal and vagus nerves. This area also contains the nuclei of the olfactory nerve.
It is a known fact that certain smells evoke certain emotions in us. So, to ensure such a reaction, the fibers of the olfactory nerve communicate with the subcortical visual analyzer, the hypothalamus and the limbic system.
Perception of olfactory information[ | ]
| External images | |
Olfactory cells[ | ]
Olfactory receptors (olfactory cells[en]) are bipolar neurons with one non-branching dendrite. It passes between the basal cells and ends in a small swelling - the olfactory club
.
Up to 20 long cilia emerge from it, which represent the sensory surface of the olfactory cell. They are usually immersed in a layer of mucus covering the epithelium and form a dense matrix with it. The olfactory cell has a dual function: perception of a stimulus and transmission of a nerve impulse to the brain, therefore it is a neurosensory cell (sensory neuron). Axons that transmit signals to the central nervous system are collected in bundles - olfactory filaments
. Olfactory neurons are capable of replacement by dividing basal cells[20][21].
The mucus in which the olfactory cilia lie contains a large number of small (20 kDa) proteins that are secreted by the nasal glands and are found in the mucus that covers not only the olfactory epithelium, but also the purely respiratory one. These proteins probably bind very non-selectively to molecules of odorants (odorants) and ensure their interaction with receptor cells[20].
Olfactory cilia do not differ in ultrastructure from other cilia and contain a regular stationary axoneme. Olfactory cilia are very long and thin: with a length of 5 to 250 microns, they reach only 100-250 in diameter. They are collected in bundles of 5-40 and emerge from the club of the olfactory cell, increasing its sensory surface. Receptor proteins are located on the surface of the cilia. Each gene from a family of genes that produces such proteins produces a certain type of them, and on the cilia of one olfactory cell there are only one type of olfactory proteins; not all genes of this family, however, can be expressed (for example, about 40% of these genes are expressed in humans). For a long time, it remained unclear whether the cilium responds to many types of odorants or only to one[11]. Now, however, it has been established that olfactory cells of one type are specific to a specific narrow class of chemical compounds, since they recognize special structural motifs in them [14] [21].
Regardless of the specificity, the sensitivity of olfactory cells is very high: they are capable of detecting substances in concentrations from 10-4 M to 10-13 M. With a cold, sensitivity decreases due to the fact that the cilia are immersed in too thick a layer of mucus [11].
In addition to the olfactory cells associated with the olfactory nerve, the nasal mucosa also contains free endings of the trigeminal nerve; they are able to react to some aggressive odors, for example, acidic or ammonia fumes[21].
Carrying out the signal[ | ]
The delivery of the olfactory stimulus begins as follows. The odorant binds to a receptor in the membrane of the olfactory cell. The olfactory receptor is a G protein coupled receptor and, like all GPCRs, contains 7 domains. Unlike other receptors of the GPCR superfamily, olfactory receptors are characterized by a large amino acid diversity in transmembrane domains[en] 3, 4 and, especially, 5. In addition, olfactory receptors differ from other GPCRs in less specificity: they have, to varying degrees, affinity to a number of stereochemically similar odorants. However, small changes in the chemical structure of the odorant may correspond to a change in the set of stimulated receptors and a change in subjective perception. Thus, replacing the hydroxyl group of octanol with a carboxyl group leads to a significant change in olfactory perception: instead of an odor reminiscent of an orange, the smell of rancidity and sweat is felt. In addition, the number of receptors stimulated and subjective perception may depend on the concentration of the odorant. For example, in low concentrations, indole has a pleasant floral aroma, and in high concentrations it has a disgusting putrefactive aroma[22].
The binding of the odorant to the receptor activates the Gs protein, which activates the enzyme adenylate cyclase, as a result of which GTP breaks down into phosphate and HDP. Adenylate cyclase converts ATP into cAMP, which binds to a cyclonucleotide-dependent cation channel in the membrane and opens the flow of Na+ and Ca2+ ions into the olfactory cell, thereby triggering an action potential in it, which is then transmitted to afferent neurons [21]. Sometimes olfactory receptors, however, activate not adenylate cyclase, but phospholipase, and the secondary messenger is not cAMP, but inositol triphosphate and diacylglycerol. In addition, it is possible that NO synthase is formed in olfactory cells due to calcium activation, which leads to the formation of cGMP [23].
Cyclonucleotide-gated channels have six hydrophobic segments and are structurally similar to voltage-gated ion channels. The difference is that cyclonucleotide-dependent channels have a large C-terminal cytoplasmic domain that binds to second messengers. There are 2400 channels/μm² on the cilia (on the olfactory club and dendrite there are only 6 channels/μm². In the absence of calcium, cyclonucleotide-dependent channels are permeable to all monovalent cations: Na+ > K+ > Li+ > Rb+ > Cs+. When exposed to an odorant, ion currents through cyclonucleotide- dependent channels change, leading to depolarization of the cell membrane and triggering an action potential[24].
Olfactory cells of the same type transmit their signals to the same glomerulus[en] of the olfactory bulb, and the spatial organization of the latter topographically repeats the location of the receptors on the surface of the olfactory concha[14]. It is worth noting that one olfactory receptor can be excited by one molecule of an odorant[25].
In 2004, Linda Buck and Richard Axel received the Nobel Prize in Physiology or Medicine for their research on mammalian olfactory receptors[26]; It was they who established the chemical nature of olfactory receptor proteins, estimated the number of genes in the mammalian genome that express these proteins, and substantiated the rules according to which one olfactory cell expresses one type of olfactory receptor proteins, and one is responsible for processing signals from all olfactory cells of the same type the same glomerulus of the olfactory bulb[27][28].
Sensory adaptation[ | ]
Interestingly, the cyclonucleotide-dependent channels of olfactory cilia do not become desensitized, that is, they do not lose sensitivity upon repeated presentation of the odorant. However, adaptation nevertheless occurs in olfactory cells. This is likely due to the entry of Ca2+ ions into the cell, which either directly or through activation of calmodulin lead to the closure of ion channels and, in addition, desensitize GPCRs[29].
In addition, the response to an olfactory stimulus is gradual, that is, a greater concentration of odorant corresponds to a greater response. This is due to the fact that cAMP increases or decreases the number of open cyclonucleotide-gated channels. To effectively distinguish signals in real time, a fast response is necessary. It has been shown that the peak of cAMP formation occurs 40-75 after exposure to an odorant and drops to zero after 100-500 ms. The G-protein cascade amplifies the signal, allowing one odorant pulse to activate multiple channels. However, the kinetics of the channels is quite slow, and the open state can lag behind the cAMP pulse by several milliseconds. With prolonged activation of GPCR odorants, cAMP pulses ensure the maintenance of cyclonucleotide-dependent channels in a constantly open state [29].
Anosmia
"Anosmia" is translated as "lack of sense of smell." If a similar condition is observed on both sides, then this indicates damage to the nasal mucosa (rhinitis, sinusitis, polyps) and, as a rule, does not threaten any serious consequences. But with unilateral loss of smell, you need to think about the fact that the olfactory nerve may be affected.
The causes of the disease may be an underdeveloped olfactory tract or fractures of the skull bones, for example, the cribriform plate. The course of the olfactory nerve is generally closely related to the bone structures of the skull. Fibers can also be damaged by bone fragments after a fracture of the nose, upper jaw, or orbit. Damage to the olfactory bulbs is also possible due to a bruise of the brain substance when falling on the back of the head.
Inflammatory diseases such as ethmoiditis, in advanced cases, melt the ethmoid bone and damage the olfactory nerve.
Olfactory pathways and information processing[ | ]
Formation of the olfactory pathways
Unlike other sensory systems (visual, auditory, somatosensory and, to a lesser extent, gustatory), where the sensory epithelium registers spatial information, the “mapping” function of the olfactory epithelium is not as pronounced. Nevertheless, it contains the rudiments of spatial organization. Mapping using 2-deoxyglucose, which makes it possible to identify active cells, showed that in the olfactory epithelium there are groups of cells associated with certain odors. Thus, butanol excites the cells of the anterior region, and limonene activates the cells of the posterior part of the mucosa. In addition, it has recently been shown that receptor cells are organized into anterior-posterior stripes (so-called expression zones
), each of which contains a complete set of cells. There appear to be 3 non-overlapping zones of expression that overlap with a smaller, fourth zone[30].
The axons of olfactory bipolar cells are united into several dozen bundles, each of which contains several hundred or thousands of fibers. They enter the cranial cavity through the openings of the ethmoid bone and unite into the olfactory nerves. The endings of the primary olfactory cells form synapses with the dendrites of the cells of the olfactory bulbs. Each such cell (mitral cell), which is a second-order sensory neuron, receives signals from about 1000 axons of primary sensory cells, that is, about 1000 olfactory axons converge on the branches of the apical dendrite of one mitral cell. About 25 such dendrites, together with terminals, form spherical formations - glomeruli
. About 2500 olfactory axons converge on one glomerulus, and in the rabbit olfactory bulb there are about 2000 glomeruli. Mitral cells are characterized by rhythmic activity caused by inhalation of odorous substances. Local interneurons of the olfactory bulbs (periglomerular cells located between the glomeruli and granular cells lying under the mitral cells) are capable of contrasting the received signals. The efferent pathways of the opposite olfactory bulb, limbic structures and reticular formation of the brain end on these cells. The system of synaptic contacts in the olfactory bulb is extremely complex, as is its chemistry: about a dozen neurotransmitters have been identified in it, including acetylcholine, dopamine, GABA and several neuropeptides[31].
The axons of the mitral cells form an olfactory tract leading to higher-order olfactory centers, which, dividing into several parts, ends on the limbic structures of the forebrain: the anterior olfactory nucleus, the septum, the piriform and parahippocampal gyri. From these structures, information enters the hippocampus, amygdala, orbitofrontal cortex (directly or through the thalamus) and the reticular formation of the midbrain[32][6].
Recognition of a specific odor is the result of the joint work of receptors and the brain, as a result of which it is represented as a combination of “primary odors.” In accordance with the Moncrieff-Eymour stereochemical theory of smell [33], humans have a seven-component odor recognition system based on the discrimination of seven
primary odors: musky, camphor, floral, ethereal, minty, pungent and putrefactive (substances belonging to the same group are similar in stereomodel)[34][35].
Anatomy of the human olfactory brain - information:
A B C D E F G H I J K L M N O P R S T U V X C CH W W E Y Z The
olfactory brain, rhinencephalon , is phylogenetically the most ancient part of the forebrain, which arose in connection with the olfactory analyzer, when the forebrain The brain has not yet become an organ of animal behavior. Therefore, all its components are different parts of the olfactory analyzer. In fish, almost the entire forebrain is the organ of smell.
With the development of the new cortex, which is observed in mammals and humans, a new part of the forebrain (neencephalon) develops - the cloak, pallium. But the cloak also goes through a long path of development and contains three parts of different phylogenetic dates.
Older parts:
- Paleopallium, part of the temporal lobe. Initially, this section was located on the lateral surface of the hemisphere, but later, under the influence of the greatly enlarging neopallium, it curled up into a sausage-shaped formation - the hippocampus and shifted medially into the cavity of the lateral ventricle of the telencephalon in the form of a protrusion of its lower horn. The hippocampus is covered with an ancient cortex, paleocortex.
- Archipallium is a small area of cortex on the ventral surface of the frontal lobe, lying near the bulbus olfactorius and covered with old cortex, archicortex.
- Neopallium, a new cloak, in the cortex of which, neocortex, higher centers of smell appeared - the cortical ends of the analyzer. This is an uncus, which is part of the vaulted gyrus. As a result, the human olfactory brain contains a number of formations of different origins, which can be topographically divided into two sections.
The peripheral section is the olfactory lobe, lobus olfactorius, which refers to a number of formations lying at the base of the brain:
- bulbus olfactorius;
- tractus olfactorius;
- trigonum olfactorium;
- substantia perforata anterior.
The central section is the convolutions of the brain:
- parahippocampal gyrus, gyrus parahippocampalis;
- dentate gyrus, gyrus dentatus;
- vaulted gyrus, gyrus fornicatus, with its anterior part located near the temporal pole - the hook, uncus.
Is something bothering you? Do you want to know more detailed information about the Olfactory Brain or do you need an examination? You can make an appointment with a doctor - the Euro lab is always at your service! The best doctors will examine you, advise you, provide the necessary assistance and make a diagnosis. You can also call a doctor at home . Euro lab clinic is open for you around the clock.
How to contact the clinic: Phone number of our clinic in Kiev: (+38 (multi-channel). The clinic secretary will select a convenient day and time for you to visit the doctor. Our coordinates and directions are listed here. Look in more detail about all the clinic’s services on its personal page.
| (+38 |
If you have previously performed any tests, be sure to take their results to a consultation with your doctor. If the studies have not been performed, we will do everything necessary in our clinic or with our colleagues in other clinics.
It is necessary to take a very careful approach to your overall health.
There are many diseases that at first do not manifest themselves in our body, but in the end it turns out that, unfortunately, it is too late to treat them.
be examined by a doctor several times a year in order not only to prevent a terrible disease, but also to maintain a healthy spirit in the body and the organism as a whole.
If you are interested in reviews about clinics and doctors, try to find the information you need on the forum.
Also register on the Euro lab to be constantly aware of the latest news and updates about the Olfactory Brain on the site, which will be automatically sent to you by email.
Other anatomical terms starting with the letter “O”:
If you are interested in any other organs and parts of the human body or have any other questions or suggestions - write to us, we will definitely try to help you
Source: https://www.eurolab-portal.ru/anatomy/376/
Anosmia[ | ]
In humans, genetic analysis has revealed several dozen specific anosmia
- disorders of the olfactory system, manifested in the inability to distinguish certain odors. For example, failure to detect the odor of cyanide occurs with a frequency of 1:10, and butyl mercaptan [en] (skunk odor) - 1:1000. Anosmia is probably caused by defects in specific olfactory receptors. Many anosmia exhibit Mendelian inheritance, but the genetics of anosmia are poorly understood[29].
Notes[ | ]
- Konstantinov, Shatalova, 2004, p. 511.
- ↑ 12
Histology, cytology and embryology, 2004, p. 340—341, 362. - Lysov, Ippolitova, Maksimov, Shevelev, 2012, p. 110-111, 575.
- Konstantinov, Naumov, Shatalova, 2012, p. 334-335.
- Vaughan, Ryan, Czaplewski, 2011, p. 27.
- ↑ 123
Silbernagl, Despopoulos, 2013, p. 352. - ↑ 12
Dzerzhinsky, Vasiliev, Malakhov, 2014, p. 389. - Histology, cytology and embryology, 2004, p. 363.
- Konstantinov, Naumov, Shatalova, 2012, p. 335.
- Plant life. Encyclopedia in 6 volumes. T. 2: Mushrooms / Ch. ed. A. A. Fedorov. - M.: Education, 1976. - 479 p. — P. 205.
- ↑ 123
Smith, 2013, p. 253. - Fleischer J., Breer H., Strotmann J. .
Mammalian Olfactory Receptors //
Frontiers in Cellular Neuroscience
, 2009,
3
(9). - P. 1-10. - doi:10.3389/neuro.03.009.2009. - Loss of olfactory capacity in primates and cetaceans (undefined)
. // Map of Life (May 29, 2008). Retrieved November 25, 2014. - ↑ 123
Assembling the Tree of Life / Ed. by J. Cracraft and M. J. Donoghue. - Oxford: Oxford University Press, 2004. - xiii + 576 p. — ISBN 0-19-517234-5. - P. 402. - Smith, 2013, p. 251.
- Meisami E., Bhatnagar KP
Structure and diversity in mammalian accessory olfactory bulb // Microscopy Research and Technique. - 1998. - Vol. 43, no. 6. - P. 476-499. - doi:10.1002/(SICI)1097-0029(19981215)43:6<476::AID-JEMT2>3.0.CO;2-V. — PMID 9880163. [] - Brennan PA, Keverne EB.
The vomeronasal organ // Handbook of Olfaction and Gustation. 2nd ed. /Ed. by R.L. Doty. - New York: Marcel Dekker, 2003. - xiv + 1121 p. — (Neurological Disease and Therapy. Book 32). — ISBN 0-8247-0719-2. — P. 967—979. - Swaney WT, Keverne EB
The evolution of pheromonal communication // Behavioral Brain Research. - 2009. - Vol. 200, no. 2. - P. 239-247. - doi:10.1016/j.bbr.2008.09.039. — PMID 18977248. [] - Smith, 2013, p. 261-262.
- ↑ 12
Smith, 2013, p. 252. - ↑ 1234
Silbernagl, Despopoulos, 2013, p. 352-353. - Smith, 2013, p. 253-254.
- Smith, 2013, p. 254.
- Smith, 2013, p. 255-256.
- Lysov, Ippolitova, Maksimov, Shevelev, 2012, p. 110.
- Axel R., Buck LB
Press Release: The 2004 Nobel Prize in Physiology or Medicine
(unspecified)
. // Nobelprize.org (October 4, 2004). Retrieved November 25, 2014. - Buck L., Axel R.
A novel multigene family may encode odorant receptors: a molecular basis for odorant recognition // Cell. - 1991. - Vol. 65, no. 1. - P. 175-187. — PMID 1840504. [] - Mombaerts P., Wang F., Dulac C., Chao SK, Nemes A., Mendelsohn M., Edmondson J., Axel R.
Visualizing an olfactory sensory map // Cell. - 1996. - Vol. 87, no. 4. - P. 675-686. — PMID 8929536. [] - ↑ 123
Smith, 2013, p. 257. - Smith, 2013, p. 251-252.
- Smith, 2013, p. 258-259.
- Tkachenko, 2009, p. 419.
- Eimour J, Johnston J, Rabin M .
Stereochemical theory of smell // Psychology of sensations and perception. 2nd ed. - M.: CheRo, 2002. - 628 p. — ISBN 5-88711-177-1. — P. 307—322. - Vorotnikov S. A.
Information devices of robotic systems. - M.: Publishing house of MSTU im. N. E. Bauman, 2005. - 384 p. — ISBN 5-7038-2207-6. - pp. 19-21. - Tkachenko, 2009, p. 417.
Literature[ | ]
- Smith K.Y.M.
Biology of sensory systems. - M.: BINOM. Knowledge Laboratory, 2013. - 583 p. — ISBN 978-5-94774-194-0. - Histology, cytology and embryology. 6th ed. / Ed. Yu. I. Afanasyeva, S. L. Kuznetsova, N. A. Yurina. - M.: Medicine, 2004. - 768 p. — ISBN 5-225-04858-7.
- Dzerzhinsky F. Ya., Vasiliev B. D., Malakhov V. V.
Zoology of vertebrates. 2nd ed. - M.: Publishing house. , 2014. - 464 p. — ISBN 978-5-4468-0459-7. - Silbernagl S., Despopoulos A.
Visual physiology. - M.: BINOM. Knowledge Laboratory, 2013. - 408 p. — ISBN 978-5-94774-385-2. - Konstantinov V.M., Naumov S.P., Shatalova S.P.
Zoology of vertebrates. 7th ed. - M.: Publishing house. , 2012. - 448 p. — ISBN 978-5-7695-9293-5. - Konstantinov V. M., Shatalova S. P.
Zoology of vertebrates. - M.: Humanitarian Publishing Center VLADOS, 2004. - 527 p. — ISBN 5-691-01293-2. - Lysov V.F., Ippolitova T.V., Maksimov V.I., Shevelev N.S.
Physiology and ethology of animals. 2nd ed. - M.: KolosS, 2012. - 605 p. — ISBN 978-5-9532-0826-0. - Tkachenko B. I., Brin V. B., Zakharov Yu. M., Nedospasov V. O., Pyatin V. F.
Human physiology. Compendium / Ed. B. I. Tkachenko. - M.: GEOTAR-Media, 2009. - 496 p. — ISBN 978-5-9704-0964-0. - Vaughan TA, Ryan JM, Czaplewski NJ
Mammalogy. 5th ed. - Sudbury, Massachusetts: Jones & Bartlett Learning, 2011. - 750 p. — ISBN 978-0-7636-6299-5.
7. Olfactory brain, its central and peripheral parts
The lower surface of the hemisphere.
The anterior sections of this surface are formed by the frontal lobe of the hemisphere, behind which the temporal pole protrudes, and there are also the lower surfaces of the temporal and occipital lobes, passing one another without noticeable boundaries.
On the lower surface of the frontal lobe, somewhat lateral and parallel to the longitudinal fissure of the cerebrum, there is an olfactory groove, sulcus
olfactorius .
Below it are the olfactory bulb and the olfactory tract, which passes from behind into the olfactory triangle, in the area of which the medial and lateral olfactory stripes are visible, striae olfactoriae medidlis et iateralis .
The area of the frontal lobe between the longitudinal fissure of the cerebrum and the olfactory sulcus is called the straight gyrus,
gyrus rectus .
The surface of the frontal lobe, lying lateral to the olfactory sulcus, is divided by deep orbital grooves,
sulci orbitdles ,
into several orbital gyri, variable in shape, location and size
, gyri orbitales .
In the posterior part of the lower surface of the hemisphere, the macollateral groove is clearly visible. Somewhat anterior to the anterior end of the collateral groove is the nasal groove,
sulcus rhlndlis .
It borders on the lateral side the curved end of the parahippocampal gyrus - the hook, uncus .
Lateral to the collateral sulcus lies
the medial occipitotemporal gyrus, gyrus occipitotetnporalis medialis .
Between this gyrus and the lateral occipitotemporal gyrus located outside of it,
gyrus occipitotetnporalis Iateralis ,
there is
an occipitotemporal sulcus, sulcus occipitotetnporalis .
The border between the lateral occipitotemporal and inferior temporal gyri is the inferolateral edge of the cerebral hemisphere.
A number of parts of the brain, located primarily on the medial surface of the hemisphere and being the substrate for the formation of such general states as wakefulness, sleep, emotions, behavioral motivation, etc., are called the “limbic system.”
Since these reactions were formed in connection with the primary functions of smell (in phylogenesis), their morphological basis is the parts of the brain that develop from the inferolateral parts of the brain bladder and belong to the so-called olfactory brain,
rhinencephalon .
The limbic system consists of the olfactory bulb, olfactory tract, olfactory triangle, anterior perforated substance, located on the lower surface of the frontal lobe (peripheral part of the olfactory brain), as well as the cingulate and parahippocampal (together with the hook) gyri, dentate gyrus, hippocampus (central part of the olfactory brain) and some other structures. Inclusion the integration of these parts of the brain into the limbic system turned out to be possible due to the common features of their structure (and origin), the presence of mutual connections and the similarity of functional reactions.