| Конечный мозг | |
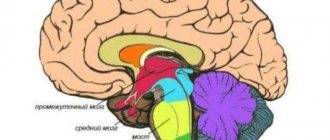
| Диаграмма, изображающая главные подотделы эмбрионального позвоночного мозга. Передний мозг в пятипузырьковой стадии разделяется на конечный мозг и промежуточный мозг | |
| Латинское название | telencephalon |
| Каталоги |
|
| Конечный мозг на Викискладе | |
Конечный мозг
(лат. telencephalon) — самый передний отдел головного мозга. Состоит из двух
полушарий большого мозга
(покрытых корой), мозолистого тела, полосатого тела и обонятельного мозга[1]. Является наиболее крупным отделом головного мозга. Это также самая развитая структура, покрывающая собой все отделы головного мозга.
Большой мозг (лат. cerebrum) состоит из двух полушарий, каждое из которых представлено корой, обонятельным мозгом и базальными ядрами. Полостью конечного мозга являются боковые желудочки, находящиеся в каждом из полушарий. Полушария большого мозга отделены друг от друга продольной щелью большого мозга и соединяются при помощи мозолистого тела, передней и задней спаек и спайки свода.
Мозолистое тело состоит из поперечных волокон, которые в латеральном направлении продолжаются в полушария, образуя лучистость мозолистого тела, соединяя друг с другом участки лобных и затылочных долей полушарий, дугообразно изгибаются и образуют передние — лобные и задние — затылочные щипцы. К задней и средней частям мозолистого тела снизу прилежит свод мозга, состоящий из двух дугообразно изогнутых тяжей, сращенных в средней своей части при помощи передней спайки мозга.
Значение обоняния для млекопитающих[ | ]
Органы обоняния млекопитающих развиты сильнее, чем у других наземных позвоночных, и играют в их жизни весьма важную роль. Способность к различению запахов млекопитающие используют для ориентирования в пространстве, при поисках пищи, в рамках межвидовых и внутривидовых контактов[4]. О важности обоняния для млекопитающих говорит и то, что наиболее обширное семейство генов[en] в их геноме образуют как раз гены, ирующие белки обонятельных рецепторов[5]. Обоняние играет определённую роль и при питании млекопитающих: приятные запахи запускают секрецию слюны и желудочного сока, а неприятные предупреждают о потенциальном вреде (например, неприятный запах испорченной пищи)[6].
По степени развития обонятельной функции млекопитающих подразделяют на две группы: макросматов
с исключительно острым обонянием (большинство млекопитающих) и
микросматов
с умеренно развитым обонянием (приматы, китообразные, ластоногие)[7].
Нос собаки — сверхчувствительный орган обоняния
Различие между данными группами хорошо видно при сопоставлении степени развития обоняния у человека и собаки. Если в носу человека имеется около 6 млн обонятельных клеток, то у собаки их — около 225 млн[8]. Многие макросматы чувствуют запахи на расстоянии нескольких сот метров и способны находить пищевые объекты под землёй[9]. Хорошо известна практика поиска растущих в лесу под землёй трюфелей при помощи специально обученных поисковых собак и свиней, которые способны учуять трюфели под землёй на расстоянии до 20 м[10].
Степень развития обоняния в целом коррелирует с числом генов, ирующих различные типы функциональных белков обонятельных рецепторов. У макросматов их обычно более 1000, у многих приматов — около 500, у человека — всего 387 (около 1—2 % генома[11]), у утконоса — 262[12]. По-видимому, хуже всего развито обоняние у китообразных; у них же наиболее высок процент псевдогенов обонятельных рецепторов[13].
Функциональная анатомия конечного мозга
Конечный мозг состоит из трех частей, возникших в разные периоды:
- Обонятельный мозг,
- Базальные ядра,
- Плащ (мантия).
Так же внутри конечного мозга — два боковых желудочка (самые большие).
Обонятельный мозг: периферический и центральны отделы
Обонятельный мозг появился с развитием органов обоняния. Делится на:
- Периферический отдел (ниже центрального):
- Обонятельные луковицы,
- Обонятельные тракты,
- Обонятельные треугольники,
- Переднее продырявленное вещество;
- Центральный отдел (на медиальной поверхности полушарий + в глубине боковых желудочков):
- Сводчатая извилина,
- Гиппокамп,
- Зубчатая извилина.
Что входит в состав базальных ядер? Их локализация
Базальные ядра — это скопление серого вещества внутри конечного мозга, окруженное белым веществом.
1) Полосатое тело – состоит из чередующихся участков серого и белого веществ.
- Хвостатое ядро,
- Чечевицеобразное ядро: скорлупа (латеральная часть),
- бледный шар (латеральные и медиальные ядра (пластинки)).
2) Ограда – форма тонкой пластинки (латерально от чечевицеобразного ядра).
3) Миндалевидное тело – базальные ядра, располагающиеся в височной доле полушарий (ближе к нижней поверхности).
Связи банальных ядер и функции
Проводящие пути базальных ядер:
- Внутренние связи – соединяют отдельные базальные ядра (одни — постоянные, другие формируются индивидуально).
- Афферентные связи (приходящие со всех отделов ГМ).
- Эфферентные связи – отходящие связи к тем отделам ГМ.
Функции базальных ядер:
- Инстинкты – безусловные рефлексы;
- Сложные автоматические движения;
- Вегетативные функции (базальные ядра входят в состав лимбической системы);
- Чувства и эмоции;
- Образование стриопаллидарной системы (согласованная работа пирамидных и экстрапирамидных систем: вначале движения контролируются пирамидной системой => затем, по мере приобретения навыков, экстрапирамидная система).
Стриопаллидарная система состоит из:
- хвостатое ядро,
- скорлупа (латеральная часть чечевицеобразного ядра),
- ограда.
- Образование паллидарной системы. Она состоит (также) из образований среднего мозга:
- бледный шар (часть чечевицеобразного ядра),
- черное вещество (средний мозг),
- красные ядра (средний мозг).
Отличие плаща от других структур конечного мозга, 3 этапа развития коры
Плащ закрывает все другие отделы конечного мозга. Состоит из серого и белого вещества.
Серое вещество – кора, представлена скоплениями тел нейронов, лежащих на поверхности (над белым веществом).
Белое вещество – проводящие пути.
Три этапа развития коры:
- Древняя кора – клетки над белым веществом — не организованы, расположены хаотично. Располагается на базальной поверхности, ближе к миндалевидному телу.
- Старая кора – появление элементов упорядоченных нервных клеток по сходству и функции. Образуются слои (не более трех).Располагается на медиальной поверхности, рядом с центральной частью обонятельного мозга.
- Новая кора – клетки строго упорядочены, различаются по форме, функции. Образуют слои (6 слоев). Клетки на разных участках — разные: Цитоархитектонические поля (около 130),
- Миелархитектонические поля (100) — различие по строению отростков,
- Нейроархитектонические поля (80) — по нейроглиям,
- Ангиоархитектонические поля — по сосудистому рисунку.
Где локализуется и чем отличается от древней и старой коры новая кора?
Новая кора покрывает 90% всей поверхности конечного мозга. Для неё характерно:
- Четкое слоистое строение (6 слоев),
- Вертикальная ориентация, импульсы идут пронизывая слои,
- Специализация каждого слоя по функциям,
- Наличие архитектонических полей.
Какие функции присущи коре? Что такое анализ и синтез?
- Аналитическая функция (анализ) – это разложение предметов и явлений на отдельные признаки;
- Синтетическая функция (синтез) – из отдельных признаков — представления о предмете или явлении.
Какие 3 морфологических субстрата обеспечивают аналитическую и синтетическую функцию?
Функции плаща:
1) Аналитическая – разложение предметов и явлений на отдельные признаки.
Для этого есть корковый центр – участок коры, способствующий восприятию одного признака.
Сама кора состоит из:
- Коркового ядра (содержит клетки, специализированные на одном признаке);
- Рассеянной части (содержит клетки, которые исполняют ощущение одного признака, но могут перестроиться на другой признак).
2) Синтетическая функция – из отдельных признаков слагается представление о предмете или явлении. Синтетические функции выполняют:
- Ассоциативные поля — зона пересечения периферических (рассеянных) частей нескольких корковых центров. Здесь — клетки, изначально способные выполнять две функции.
- Ассоциативные нервные волокна.
Где локализуются центры: слуха, зрения, вкуса, обоняния, кожного чувства, двигательного анализатора, равновесия?
Центры первой сигнальной системы (система биологической сущности человека, биологические функции которых имеются у других животных).
- Центр слуха – верхневисочная извилина,
- Центр кожного чувства (тактильного) чувства – верхнетеменная долька,
- Температурное и болевое чувство – задняя постцентральная извилина,
- Центр стереогнозии – парацентральная долька,
- Проприоцептивное чувство – передняя центральная извилина (3 слой коры),
- Центр зрения – по берегам шпорной борозды (борозды птичьей шпоры),
- Центр вкуса и обоняния – крючок,
- Центр двигательного анализатора – передняя центральная извилина (5 слой коры),
- Центр практических навыков – в области надкраевой извилины.
Локализация центров второй сигнальной системы
Вторая сигнальная система – обеспечение высшей нервной деятельности человека. Все центры сводят к речи человека:
- Центр артикуляции речи – в задней части нижней лобной извилины;
- Центр письма и письменной речи – в задней части средней лобной извилины;
- Центр восприятия устной речи – верхний отдел верхней височной извилины;
- Центр чтения и восприятие письменной речи – угловая извилина.
Центры второй сигнальной системы располагаются рядом со сходными по функции первой сигнальной системы.
Интегративная функция коры: на чем основана, чем обеспечивается? Куда идут нисходящие пути от коры?
Кора полушарий объединяет всю нервную систему в единое целое, а через нервную систему — весь организм.
Интегративная функция основана на проекционных проводящих путях. Они связывают конечный мозг с нижележащими отделами ГМ. Это осуществляется через сложную рефлекторную дугу. Включает два образования:
- Внутренняя капсула – включает все проекционные пути;
- Лучистый венец.
Что относится к лимбической системе?
- Центральная часть обонятельного мозга,
- Миндалевидный комплекс,
- Прозрачная перегородка,
- Свод,
- Гиппокамп,
- Орбитальная кора.
Все это – конечный мозг.
Связи лимбической системе
- Внутренние (короткие) связи – проходят по двум кругам Папеца( большой, малый круг)
Импульсы по внутренним связям лимбической системы стекаются в гиппокамп и миндалевидный комплекс – это сердцевина лимбической системы.
- Внешние (длинные) связи:
- Двусторонние;
- Связь со всеми корковыми центрами;
- Со всеми ассоциативными полями коры (интегративные участки коры);
- Двусторонняя связь с гипоталамусом, ретикулярной формацией, стволом головного мозга.
Кроме того, ЛС получает коллатерали от всех чувствительных путей.
Функции лимбической системы
- Инстинктивное поведение (у человека инстинкты контролируются корой),
- Вегетативная функция (висцеральный мозг),
- Эндокринная функция,
- Сон и бодрствование,
- Память (долгосрочная и краткосрочная),
- Внимание (активное и пассивное),
- Эмоциональный статус,
- Определение пола человека (миндалевидный комплекс).
3 0
Источник: https://medfsh.ru/teoriya/teoriya-po-anatomii/voprosy-po-anatomii/funktsionalnaya-anatomiya-konechnogo-mozga
Органы обоняния[ | ]
Обонятельная система млекопитающих на примере человека: 1 — обонятельная луковица, 2 — митральные клетки[en], 3 — кость, 4 — носовой эпителий, 5 — обонятельный клубочек[en], 6 — обонятельные рецепторы
Органы обоняния млекопитающих располагаются в задней верхней части носовой полости, где возникает весьма сложная, особенно у макросматов, система обонятельных носовых раковин[en]
— тонких костных лепестков, направленных внутрь полости и покрытых обонятельным эпителием. В носовых раковинах не только происходит обонятельный анализ вдыхаемого воздуха, но и его нагревание перед поступлением в лёгкие. Среди современных видов четвероногих обонятельные раковины имеются лишь у млекопитающих, а также у немногих видов птиц, у которых эти раковины возникли независимо[14]. Обонятельный эпителий содержит обонятельные рецепторные клетки, опорные клетки, секретирующие слизь и по свойствам близкие к глиальным клеткам, а также базальные клетки, которые, подобно стволовым клеткам, способны делиться и давать начало новым функциональным нейронам в течение всей жизни животного. Размеры обонятельного эпителия у млекопитающих варьируют от 2—4 см² (человек) и 9,3 см² (кролик) до 18 см² (собака) и 21 см² (домашняя кошка). Впрочем, эти значения не дают представления об остроте обоняния, потому что в них не учитывается количество обонятельных рецепторов на единицу поверхности. Обонятельные рецепторы способны улавливать содержащиеся во вдыхаемом воздухе молекулы пахучих веществ. Как и рецепторы вкуса, их относят к группе хеморецепторов. Сигналы о наличии пахнущих веществ передаются через обонятельный нерв в соответствующий центр головного мозга — обонятельную луковицу, или первичные центры обоняния коры головного мозга. Из последней обонятельные сигналы передаются в гипоталамус, лимбическую систему, ретикулярную формацию и неокортекс[7][6][15].
Большинство млекопитающих сохраняют якобсонов орган как обособленный отдел обонятельной капсулы. Этот орган, имеющийся также у двояышащих и большинства четвероногих (важнейшие исключения — птицы и кроилы), служит главным образом для восприятия феромонов. У представителей ряда групп (китообразные, сирены, большинство рукокрылых и узконосые приматы, включая человека) якобсонов орган рудиментарен или полностью утрачен[16][17][18].
Вомероназальный орган выстлан обонятельным эпителием, похожим на тот, что покрывает носовые раковины носа. Обонятельные рецепторные клетки возобновляются в течение всей жизни и поддерживаются эпителиальными и базальными клетками, однако вместо ресничек имеют микроворсинки (микровилли). Рецепторные молекулы также представлены GPCR, однако их аминокислотная последовательность не имеет ничего общего с рецепторами носовых раковин. GPCR вомероназального органа представлены двумя различными семействами, каждое из которых содержит от 100 до 200 генов и развивалось независимо. Представители одного из этих семейств имеют длинный внеклеточный N-концевой домен, сходный с метаботропным глутаматным рецептором. Вторичным посредником здесь служит не цАМФ, как в раковинах носа, а инозитолтрифосфат. Афферентные волокна из вомероназального эпителия проецируются в дополнительную обонятельную луковицу, которая у большинства расположена кзади от основной обонятельной луковицы. Как и обонятельный эпителий носа, вомероназальный эпителий тоже подразделяется на зоны: в апикальной и базальной частях органа экспрессируются разные G-белки. Эти зоны сохраняются и в проекциях в дополнительную обонятельную луковицу: апикальная зона эпителия проецируется в переднюю зону луковицы, а базальная — в заднюю. Гломерулы в дополнительной луковице выражены хуже, чем в основной. Кроме того, вместо пространственной карты основной луковицы представительство дополнительной луковицы более сложное и мозаичное. Дополнительная луковица не имеет проекций в кору и связана только с лимбической системой: с миндалиной и гипоталамическими ядрами, которые играют важную роль в половом поведении. Возможно, дополнительная луковица реагирует только на особые видоспецифические сочетания веществ соответствующего феромона и просто игнорирует все остальные[19].
Функции
Само название структуры указывает на то, для чего она предназначена. Функции обонятельного нерва заключаются в улавливании запаха и его расшифровке. Они вызывают аппетит и слюноотделение, если аромат приятный, или, напротив, провоцируют тошному и рвоту, когда амбре оставляет желать лучшего.
Для того чтобы достичь такого эффекта, обонятельный нерв проходит через ретикулярную формацию и направляется в ствол мозга. Там волокна соединяются с ядрами промежуточного, языкоглоточного и блуждающего нервов. В этой области так же находятся ядра обонятельного нерва.
Известны факт, что те или иные запахи вызывают у нас определенные эмоции. Так вот, чтобы обеспечить подобную реакцию, волокна обонятельного нерва связываются с подкорковым зрительным анализатором, гипоталамусом и лимбической системой.
Восприятие обонятельной информации[ | ]
| Внешние изображения | |
Обонятельные клетки[ | ]
Обонятельные рецепторы (обонятельные клетки[en]) представляют собой биполярные нейроны с одним неветвящимся дендритом. Он проходит между базальными клетками и оканчивается небольшим вздутием — обонятельной булавой
. Из неё выходят до 20 длинных ресничек, которые представляют собой сенсорную поверхность обонятельной клетки. Они, как правило, погружены в слой слизи, покрывающей эпителий, и образуют с ним плотный матрикс. Обонятельная клетка имеет двойную функцию: восприятие стимула и передача нервного импульса в мозг, поэтому является нейросенсорной клеткой (сенсорным нейроном). Аксоны, выполняющие передачу сигналов в ЦНС, собраны в пучки —
обонятельные нити
. Обонятельные нейроны способны к замещению путём деления базальных клеток[20][21].
Слизь, в которой лежат обонятельные реснички содержит большое количество некрупных (20 кДа) белков, которые секретируются железами носа и встречаются в слизи, покрывающей не только обонятельный эпителий, но и чисто респираторный. Эти белки, возможно, очень неселективно связываются с молекулами пахучих веществ (одорантами) и обеспечивают их взаимодействие с рецепторными клетками[20].
Обонятельные реснички по ультраструктуре не отличаются от других ресничек и содержат обычную неподвижную аксонему. Обонятельные реснички очень длинные и тонкие: при длине от 5 до 250 мкм они достигают лишь 100—250 в диаметре. Они собраны в пучки по 5—40 и выходят из булавы обонятельной клетки, увеличивая её сенсорную поверхность. Рецепторные белки располагаются на поверхности ресничек. Каждый ген из семейства генов, ирующих такие белки, ирует определённую их разновидность, и на ресничках одной обонятельной клетки присутствуют обонятельные белки лишь одной разновидности; не все гены данного семейства, впрочем, могут экспрессироваться (так, у человека экспрессируется около 40 % этих генов). Долгое время оставалось неясным, отвечает ли ресничка на многие типы одорантов или только на один[11]. Сейчас, однако, установлено, что обонятельные клетки одного типа специфичны к конкретному узкому классу химических соединений, поскольку распознают в них особые структурные мотивы[14][21].
Независимо от специфичности, чувствительность обонятельных клеток очень велика: они способны регистрировать вещества в концентрации от 10-4 М до 10-13 М. При простуде чувствительность снижается из-за того, что реснички оказываются погружёнными в слишком толстый слой слизи[11].
Кроме обонятельных клеток, связанных с обонятельным нервом, в слизистой оболочке носа имеются также свободные окончания тройничного нерва; они способны реагировать на некоторые агрессивные запахи, например, кислотные или аммиачные испарения[21].
Проведение сигнала[ | ]
Проведение обонятельного стимула начинается следующим образом. Пахучее вещество связывается с рецептором в мембране обонятельной клетки. Обонятельный рецептор представляет собой рецептор, связанный с G-белком (англ. G protein coupled receptor) и, как и все GPCR, содержит 7 доменов. В отличие от других рецепторов надсемейства GPCR, для обонятельных рецепторов характерно большое аминокислотное разнообразие в трансмембранных доменах[en] 3, 4 и, особенно, 5. Кроме того, от других GPCR обонятельные рецепторы отличаются меньшей специфичностью: они в той или иной степени имеют сродство к целому ряду стереохимически сходных одорантов. Однако небольшим изменениям химической структуры одоранта может соответствовать изменение набора стимулируемых рецепторов и изменение субъективного восприятия. Так, замещение гидроксильной группы октанола на карбоксильную приводит к значительному изменению обонятельного восприятия: вместо запаха, напоминающего запах апельсина, ощущается запах прогорклости и пота. Кроме того, количество стимулируемых рецепторов и субъективное восприятие может зависеть от концентрации одоранта. Например, в низкой концентрации индол имеет приятный цветочный аромат, а в высокой — отвратительный гнилостный[22].
Связывание одоранта с рецептором активирует Gs-белок[en], который активирует фермент аденилатциклазу, в результате чего ГТФ распадается на фосфат и ГДФ. Аденилатциклаза переводит АТФ в цАМФ, который связывается с циклонуклеотид-зависимым катионным каналом в мембране и открывает ток ионов Na+ и Са2+ в обонятельную клетку, тем самым запуская потенциал действия в ней, который затем передается на афферентные нейроны[21]. Иногда обонятельные рецепторы, правда, активируют не аденилатциклазу, а фосфолипазу, и вторичным посредником выступает не цАМФ, а инозитолтрифосфат и диацилглицерин. Кроме того, возможно, что в обонятельных клетках из-за активации кальцием NO-синтазы образуется , который приводит к образованию цГМФ[23].
Циклонуклеотид-зависимые каналы имеют шесть гидрофобных сегментов и по структуре напоминают потенциал-зависимые ионные каналы. Отличие заключается в наличии у циклонуклеотид-зависимых каналов большого С-концевого цитоплазматического домена, который связывается со вторичными посредниками. На ресничках расположено 2400 каналов/мкм² (на обонятельной булаве и дендрите лишь 6 каналов/мкм². В отсутствие кальция циклонуклеотид-зависимые каналы проницаемы для всех одновалентных катионов: Na+ > K+ > Li+ > Rb+ > Cs+. При воздействии одоранта ионные токи через циклонуклеотид-зависимые каналы изменяются, приводя к деполяризации мембраны клетки и запуску потенциала действия[24].
Обонятельные клетки одного типа передают свои сигналы в один и тот же клубочек[en] обонятельной луковицы, и пространственная организация последней топографически повторяет расположение рецепторов на поверхности обонятельной раковины[14]. Стоит отметить, что один обонятельный рецептор может быть возбуждён одной молекулой пахучего вещества[25].
В 2004 году Линда Бак и Ричард Эксел получили Нобелевскую премию в области физиологии и медицины за исследования обонятельных рецепторов млекопитающих[26]; именно они установили химическую природу белков обонятельных рецепторов, дали оценку числа генов в геноме млекопитающих, ирующих эти белки, и обосновали правила, по которым одна обонятельная клетка экспрессирует одну разновидность белков обонятельных рецепторов, а за обработку сигналов всех обонятельных клеток одного типа отвечает один и тот же клубочек обонятельной луковицы[27][28].
Сенсорная адаптация[ | ]
Интересно, что циклонуклеотид-зависимые каналы обонятельных ресничек не десенситизируются[en], т. е. не утрачивают чувствительности при повторных предъявлениях одоранта. Однако в обонятельных клетках, тем не менее, происходит адаптация. Вероятно, это происходит из-за входа в клетку ионов Ca2+, которые или непосредственно, или через активацию кальмодулина приводят к закрытию ионных каналов и, кроме того, десенситизируют GPCR[29].
Кроме того, ответ на обонятельный стимул градуален, то есть большей концентрации одоранта соответствует больший ответ. Это связано с тем, что цАМФ увеличивает или уменьшает количество открытых циклонуклеотид-зависимые каналов. Для эффективного различения сигналов в реальном времени необходим быстрый ответ. Показано, что пик образования цАМФ наступает через 40—75 после воздействия пахучего вещества и через 100—500 мс падает до нуля. G-белковый каскад усиливает сигнал, благодаря чему один импульс одоранта активирует множество каналов. Впрочем, кинетика каналов достаточно медленна, и открытое состояние может отставать от импульса цАМФ на несколько миллисекунд. При продолжительной активации GPCR одорантов импульсы цАМФ обеспечивают поддержание циклонуклеотид-зависимые каналов в постоянно открытом состоянии[29].
Аносмия
«Аносмия» переводится как «отсутствие обоняния». Если подобное состояние наблюдается с двух сторон, то это свидетельствует в пользу поражения слизистой носа (риниты, синуситы, полипы) и, как правило, не грозит никакими серьезными последствиями. Но при односторонней утрате обоняния необходимо задуматься о том, что может быть поражен обонятельный нерв.
Причинами заболевания могжет быть недоразвитый обонятельный тракт или переломы костей черепа, например, решетчатой пластинки. Ход обонятельного нерва вообще тесно связан с костными структурами черепа. Повредить волокна могут и осколки кости после перелома носа, верхней челюсти, глазницы. Повреждение обонятельных луковиц возможно и из-за ушиба вещества мозга, при падении на затылок.
Воспалительные заболевания, такие как этмоидит, в запущенных случаях расплавляют решетчатую кость и повреждают обонятельный нерв.
Обонятельные пути и обработка информации[ | ]
Формирование обонятельных путей
В отличие от других сенсорных систем (зрительной, слуховой, соматосенсорной и, в меньшей степени, вкусовой), где сенсорный эпителий регистрирует пространственную информацию, «картирующая» функция обонятельного эпителия не так выражена. Тем не менее, в нём присутствуют зачатки пространственной организации. Картирование при помощи 2-дезоксиглюкозы[en], которая даёт возможность выявить активные клетки, показало, что в обонятельном эпителии имеются группы клеток, связанные с определёнными запахами. Так, бутанол возбуждает клетки передней области, а лимонен активирует клетки задней части слизистой. Кроме того, недавно было показано, что рецепторные клетки организованы в передне-задние полосы (так называемые зоны экспрессии
), каждая из которых содержит полный набор клеток. Судя по всему, существует 3 неперекрывающиеся зоны экспрессии, которые перекрываются с меньшей, четвёртой зоной[30].
Аксоны обонятельных биполярных клеток объединяются в несколько десятков пучков, каждый из которых содержит несколько сотен или тысяч волокон. Они входят в полость черепа через отверстия решётчатой кости и объединяются в обонятельные нервы. Окончания первичных обонятельных клеток образуют синапсы с дендритами клеток обонятельных луковиц. Каждая такая клетка (митральная клетка[en]), являющаяся сенсорным нейроном второго порядка, получает сигналы от около 1000 аксонов первичных сенсорных клеток, то есть около 1000 обонятельных аксонов конвергируют на разветвлениях апикального дендрита одной митральной клетки. Около 25 таких дендритов совместно с терминалями формируют сферические образования — гломерулы
. На одной гломеруле конвергирует около 2500 обонятельных аксонов, а в обонятельной луковице кролика имеется около 2000 гломерул. Для митральных клеток характерна ритмическая активность, обусловленная вдыханием пахучих веществ. Локальные интернейроны обонятельных луковиц (перигломерулярные клетки, расположенные между гломерулами, и зернистые клетки, залегающие под митральными клетками) способны к контрастированию получаемых сигналов. На этих клетках оканчиваются эфферентные[en] пути противоположной обонятельной луковицы, лимбических структур и ретикулярной формации мозга. Система синаптических контактов в обонятельной луковицы чрезвычайно сложна, как и её химия: в ней идентифицировано около дюжины нейромедиаторов, среди которых ацетилхолин, дофамин, ГАМК и несколько нейропептидов[31].
Аксоны митральных клеток образуют обонятельный тракт, ведущий к обонятельным центрам высшего порядка, который, разделяясь на несколько частей, оканчивается на лимбических структурах переднего мозга: переднем обонятельном ядре[en], перегородке, пириформной и парагиппокампальной извилинах. От этих структур информация поступает в гиппокамп, миндалины, орбитофронтальную кору (напрямую или через таламус) и ретикулярную формацию среднего мозга[32][6].
Распознавание конкретного запаха является результатом совместной работы рецепторов и мозга, в результате чего он представляется как комбинация «первичных запахов». В соответствии со стереохимической теорией обоняния Монкриффа — Эймура[33], у человека имеется семикомпонентная система распознавания запахов, базирующаяся на различении семи
первичных запахов: мускусного, камфарного, цветочного, эфирного, мятного, едкого и гнилостного (относящиеся к одной группе вещества сходны в стереомодели)[34][35].
Анатомия Обонятельнего мозга человека – информация:
А Б В Г Д Е Ж З И К Л М Н О П Р С Т У Ф Х Ц Ч Ш Щ Э Ю Я
Обонятельный мозг, rhinencephalon, есть филогенетически самая древняя часть переднего мозга, возникшая в связи с анализатором обоняния, когда передний мозг не стал еще органом поведения животного. Поэтому все компоненты его являются различными частями обонятельного анализатора. У рыб почти весь передний мозг является органом обоняния.
С развитием новой коры, что наблюдается у млекопитающих и человека, развивается новая часть переднего мозга (neencephalon) – плащ, pallium. Но и плащ проходит свой длинный путь развития и содержит три части различной филогенетической давности.
Более старые части:
- Paleopallium, входящий в состав височной доли. Вначале этот отдел располагался на латеральной поверхности полушария, но в дальнейшем, под влиянием сильно увеличивающегося neopallium, он свернулся в колбасовидное образование – гиппокамп и сместился медиально в полость бокового желудочка конечного мозга в виде выпячивания его нижнего рога. Гиппокамп покрыт древней корой, paleocortex.
- Archipallium – небольшой участок коры на вентральной поверхности лобной доли, лежащий вблизи bulbus olfactorius и покрытый старой корой, archicortex.
- Neopallium, новый плащ, в коре которого, neocortex, появились высшие центры обоняния – корковые концы анализатора. Это – uncus, являющийся частью сводчатой извилины. В результате обонятельный мозг человека содержит ряд образований различного происхождения, которые топографически можно разделить на два отдела.
Периферический отдел – это обонятельная доля, lobus olfactorius, под которой разумеется ряд образований, лежащих на основании мозга:
- bulbus olfactorius;
- tractus olfactorius;
- trigonum olfactorium;
- substantia perforata anterior.
Центральный отдел – это извилины мозга:
- парагиппокампальная извилина, gyrus parahippocampalis;
- зубчатая извилина, gyrus dentatus;
- сводчатая извилина, gyrus fornicatus, с расположенной вблизи височного полюса передней ее частью – крючком, uncus.
Вас что-то беспокоит? Вы хотите узнать более детальную информацию о Обонятельном мозге или же Вам необходим осмотр? Вы можете записаться на прием к доктору – клиника Eurolab всегда к Вашим услугам! Лучшие врачи осмотрят Вас, проконсультируют, окажут необходимую помощь и поставят диагноз. Вы также можете вызвать врача на дом. Клиника Eurolab открыта для Вас круглосуточно.
Как обратиться в клинику: Телефон нашей клиники в Киеве: (+38 (многоканальный). Секретарь клиники подберет Вам удобный день и час визита к врачу. Наши координаты и схема проезда указаны здесь. Посмотрите детальнее о всех услугах клиники на ее персональной странице.
| (+38 |
Если Вами ранее были выполнены какие-либо исследования, обязательно возьмите их результаты на консультацию к врачу. Если исследования выполнены не были, мы сделаем все необходимое в нашей клинике или у наших коллег в других клиниках.
Необходимо очень тщательно подходить к состоянию Вашего здоровья в целом.
Есть много болезней, которые по началу никак не проявляют себя в нашем организме, но в итоге оказывается, что, к сожалению, их уже лечить слишком поздно.
Для этого просто необходимо по несколько раз в год проходить обследование у врача, чтобы не только предотвратить страшную болезнь, но и поддерживать здоровый дух в теле и организме в целом.
Если Вас интересуют отзывы о клиниках и врачах – попробуйте найти нужную Вам информацию на форуме.
Также зарегистрируйтесь на медицинском портале Eurolab, чтобы быть постоянно в курсе последних новостей и обновлений информации о Обонятельном мозге на сайте, которые будут автоматически высылаться Вам на почту.
Другие анатомические термины на букву “О”:
Если Вас интересуют еще какие-нибудь органы и части тела человека или у Вас есть какие-либо другие вопросы или предложения – напишите нам, мы обязательно постараемся Вам помочь
Источник: https://www.eurolab-portal.ru/anatomy/376/
Аносмии[ | ]
У человека генетический анализ выявил несколько дюжин специфических аносмий
— расстройств обонятельной системы, проявляющихся в неспособности различать определённые запахи. Например, неспособность определить запах цианида встречается с частотой 1:10, а бутилмеркаптана[en] (пахучее вещество скунса) — 1:1000. Вероятно, аносмии обусловлены дефектами специфических обонятельных рецепторов. Многие аносмии демонстрируют менделевское наследование, однако генетика аносмий изучена плохо[29].
Примечания[ | ]
- Константинов, Шаталова, 2004, с. 511.
- ↑ 12
Гистология, цитология и эмбриология, 2004, с. 340—341, 362. - Лысов, Ипполитова, Максимов, Шевелёв, 2012, с. 110—111, 575.
- Константинов, Наумов, Шаталова, 2012, с. 334—335.
- Vaughan, Ryan, Czaplewski, 2011, p. 27.
- ↑ 123
Зильбернагль, Деспопулос, 2013, с. 352. - ↑ 12
Дзержинский, Васильев, Малахов, 2014, с. 389. - Гистология, цитология и эмбриология, 2004, с. 363.
- Константинов, Наумов, Шаталова, 2012, с. 335.
- Жизнь растений. Энциклопедия в 6 тт. Т. 2: Грибы / Гл. ред. А. А. Фёдоров. — М.: Просвещение, 1976. — 479 с. — С. 205.
- ↑ 123
Смит, 2013, с. 253. - Fleischer J., Breer H., Strotmann J. .
Mammalian Olfactory Receptors //
Frontiers in Cellular Neuroscience
, 2009,
3
(9). — P. 1—10. — doi:10.3389/neuro.03.009.2009. - Loss of olfactory capacity in primates and cetaceans (неопр.)
. // Map of Life (29 мая 2008). Дата обращения 25 ноября 2014. - ↑ 123
Assembling the Tree of Life / Ed. by J. Cracraft and M. J. Donoghue. — Oxford: Oxford University Press, 2004. — xiii + 576 p. — ISBN 0-19-517234-5. — P. 402. - Смит, 2013, с. 251.
- Meisami E., Bhatnagar K. P.
Structure and diversity in mammalian accessory olfactory bulb // Microscopy Research and Technique. — 1998. — Vol. 43, no. 6. — P. 476—499. — doi:10.1002/(SICI)1097-0029(19981215)43:6<476::AID-JEMT2>3.0.CO;2-V. — PMID 9880163. [] - Brennan P. A., Keverne E. B. .
The vomeronasal organ // Handbook of Olfaction and Gustation. 2nd ed. / Ed. by R. L. Doty. — New York: Marcel Dekker, 2003. — xiv + 1121 p. — (Neurological Disease and Therapy. Book 32). — ISBN 0-8247-0719-2. — P. 967—979. - Swaney W. T., Keverne E. B.
The evolution of pheromonal communication // Behavioural Brain Research. — 2009. — Vol. 200, no. 2. — P. 239—247. — doi:10.1016/j.bbr.2008.09.039. — PMID 18977248. [] - Смит, 2013, с. 261—262.
- ↑ 12
Смит, 2013, с. 252. - ↑ 1234
Зильбернагль, Деспопулос, 2013, с. 352—353. - Смит, 2013, с. 253—254.
- Смит, 2013, с. 254.
- Смит, 2013, с. 255—256.
- Лысов, Ипполитова, Максимов, Шевелёв, 2012, с. 110.
- Axel R., Buck L. B.
Press Release: The 2004 Nobel Prize in Physiology or Medicine
(неопр.)
. // Nobelprize.org (4 октября 2004). Дата обращения 25 ноября 2014. - Buck L., Axel R.
A novel multigene family may encode odorant receptors: a molecular basis for odor recognition // Cell. — 1991. — Vol. 65, no. 1. — P. 175—187. — PMID 1840504. [] - Mombaerts P., Wang F., Dulac C., Chao S. K., Nemes A., Mendelsohn M., Edmondson J., Axel R.
Visualizing an olfactory sensory map // Cell. — 1996. — Vol. 87, no. 4. — P. 675—686. — PMID 8929536. [] - ↑ 123
Смит, 2013, с. 257. - Смит, 2013, с. 251—252.
- Смит, 2013, с. 258—259.
- Ткаченко, 2009, с. 419.
- Эймур Дж., Джонстон Дж., Рабин М. .
Стереохимическая теория обоняния // Психология ощущений и восприятия. 2-е изд. — М.: ЧеРо, 2002. — 628 с. — ISBN 5-88711-177-1. — С. 307—322. - Воротников С. А. .
Информационные устройства робототехнических систем. — М.: Изд-во МГТУ им. Н. Э. Баумана, 2005. — 384 с. — ISBN 5-7038-2207-6. — С. 19—21. - Ткаченко, 2009, с. 417.
Литература[ | ]
- Смит К. Ю. М. .
Биология сенсорных систем. — М.: БИНОМ. Лаборатория знаний, 2013. — 583 с. — ISBN 978-5-94774-194-0. - Гистология, цитология и эмбриология. 6-е изд / Под ред. Ю. И. Афанасьева, С. Л. Кузнецова, H. А. Юриной. — М.: Медицина, 2004. — 768 с. — ISBN 5-225-04858-7.
- Дзержинский Ф. Я., Васильев Б. Д., Малахов В. В.
Зоология позвоночных. 2-е изд. — М.: Издат. , 2014. — 464 с. — ISBN 978-5-4468-0459-7. - Зильбернагль С., Деспопулос А.
Наглядная физиология. — М.: БИНОМ. Лаборатория знаний, 2013. — 408 с. — ISBN 978-5-94774-385-2. - Константинов В. М., Наумов С. П., Шаталова С. П.
Зоология позвоночных. 7-е изд. — М.: Издат. , 2012. — 448 с. — ISBN 978-5-7695-9293-5. - Константинов В. М., Шаталова С. П.
Зоология позвоночных. — М.: Гуманитарный издательский центр ВЛАДОС, 2004. — 527 с. — ISBN 5-691-01293-2. - Лысов В. Ф., Ипполитова Т. В., Максимов В. И., Шевелёв Н. С.
Физиология и этология животных. 2-е изд. — М.: КолосС, 2012. — 605 с. — ISBN 978-5-9532-0826-0. - Ткаченко Б. И., Брин В. Б., Захаров Ю. М., Недоспасов В. О., Пятин В. Ф.
Физиология человека. Compendium / Под ред. Б. И. Ткаченко. — М.: ГЭОТАР-Медиа, 2009. — 496 с. — ISBN 978-5-9704-0964-0. - Vaughan T. A., Ryan J. M., Czaplewski N. J.
Mammalogy. 5th ed. — Sudbury, Massachusetts: Jones & Bartlett Learning, 2011. — 750 p. — ISBN 978-0-7636-6299-5.
7.Обонятельный мозг, его центральный и периферический отделы
Нижняяповерхность полушария.
Передниеотделы этой поверхности образованылобной долей полушария, позади которойвыступает височный полюс, а такженаходятся нижние поверхности височнойи затылочной долей, переходящие одна вдругую без заметных границ.
Нанижней поверхности лобной доли, нескольколатеральнее и параллельно продольнойщели большого мозга, находитсяобонятельнаяборозда,
sulcusolfactorius.
Снизук ней прилежат обонятельная луковицаи обонятельный тракт, переходящий сзадив обонятельный треугольник, в областикоторого видны медиальная и латеральнаяобонятельные полоски, striaeolfactoriaemedidlisetiateralis.
Участоклобной доли между продольной щельюбольшого мозга и обонятельной бороздойполучил название прямойизвилины,
gyrusrectus.
Поверхностьлобной доли, лежащая латеральнееобонятельной борозды, разделенанеглубокими глазничнымибороздами,
sulciorbitdles,
нанесколько вариабельных по форме,расположению и размерам
глазничныхизвилин,gyriorbitales.
Взаднем отделе нижней поверхностиполушария хорошо различимаколлатеральная борозда. Несколькокпереди от переднего конца коллатеральнойборозды находится носоваяборозда,
sulcusrhlndlis.
Онаограничивает с латеральной стороныизогнутый конец парагиппокампальнойизвилины — крючок, uncus.
Латеральнееколлатеральной борозды лежит
медиальнаязатылочно-височная извилина,gyrusoccipitotetnporalismedialis.
Междуэтой извилиной и расположеннойкнаружи от нее латеральнойзатылочно-височной извилиной,
gyrusoccipitotetnporalisIateralis,
находится
затылочно-височнаяборозда,sulcusoccipitotetnporalis.
Границеймежду латеральной затылочно-височнойи нижней височной извилинами служитнижнелатеральный край полушариябольшого мозга.
Рядотделов головного мозга, расположенныхпреимущественно на медиальнойповерхности полушария и являющихсясубстратом для формирования такихобщих состояний, как бодрствование,сон, эмоции, мотивации поведения и др.,выделяют под названием «лимбическаясистема».
Поскольку эти реакциисформировались в связи с первичнымифункциями обоняния (в филогенезе),их морфологической основой являются отделы мозга, которые развиваются изнижнелатеральных отделов мозговогопузыря и относятся к так называемомуобонятельномумозгу,
rhinencephalon.
Лимби-ческуюсистему составляют обонятельнаялуковица, обонятельный тракт, обонятельныйтреугольник, переднее продырявленноевещество, расположенные на нижнейповерхности лобной доли (периферическийотдел обонятельного мозга), а такжепоясная и парагиппокампальная (вместес крючком) извилины, зубчатая извилина,гиппокамп (центральный отдел обонятельногомозга) и некоторые другие структуры.Включение этих отделов мозга в лимбическуюсистему оказалось возможным в связи собщими чертами их строения (и происхождения),наличием взаимных связей и сходством функциональных реакций.