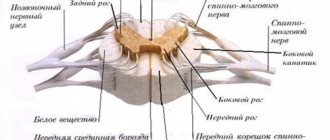
Расположение
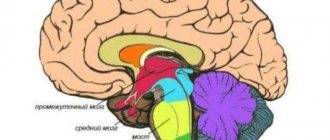
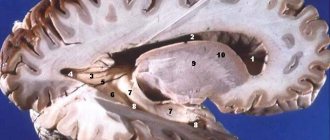
Таламус является частью больших полушарий переднего мозга. Он расположен латеральней боковых желудочков — полостей мозга, которые являются частью системы циркуляции ликвора (спинномозговой жидкости). Снизу от него находится гипоталамус, от которого зрительные бугры отделены бороздой.
Выше и несколько снаружи от таламуса находятся базальные ядра. Эти образования необходимы для осуществления точных, координированных движений. Друг от друга эти структуры разделены внутренней капсулой — пучком белого вещества переднего мозга, через который проходят проводящие пути от периферии к центру.
Между собой правая и левая части таламуса соединены межталамическим серым веществом. Оно присутствует у 70% людей.
Классификация ядер таламуса
Всего в зрительных буграх мозга насчитывается около 120 ядер. В зависимости от места нахождения их подразделяют на три группы:
- медиальные;
- латеральные;
- передние.
В латеральной группе ядер выделяют, в свою очередь, медиальное и латеральное коленчатые тела, а также подушку.
Существует также классификация в зависимости от выполняемой функции ядрами:
- специфические;
- ассоциативные;
- неспецифические.
Сравнительная анатомия
Сравнительно-анатомические и эмбриологические исследования выявили весьма сложный характер развития Таламуса и отдельных его комплексов, определяемый как общими закономерностями эволюции мозга хордовых, так и экологическими факторами формирования сенсорных систем у различных представителей этого типа.
Образования вентрального Т., особенно наиболее древняя вентральная часть латерального коленчатого тела, дифференцируются уже у селахий, т. е. значительно раньше структур дорсального Т., подразделение к-рого на отдельные образования выявляется лишь у рептилий. У птиц и рептилий наиболее развитой структурой дорсального Т. является круглое ядро, не имеющее прямых гомологов в дорсальном Т. млекопитающих. Наиболее сложно дифференцированы структуры дорсального Т. у млекопитающих. Древнейшие образования дорсального Т.— заднее и претектальное ядра, хорошо развитые у низших млекопитающих, у высших — редуцируются. Подобный процесс происходит и с интраламинарными (внутрипластинчатыми, Т.) ядрами, а также с вентральной частью ядра латерального коленчатого тела, относящимся к вентральному Т.
Усложнение дифференцировки структур дорсального Т. связано с еще большим усложнением строения новой коры (неокортекса) мозга млекопитающих, где закономерности дивергентного (разнонаправленного) развития проявляются в относительном прогрессировании новейших и относительном регрессировании филогенетически древнейших зон коры полушарий головного мозга. Этот процесс объясняется тем, что по мере эволюционного усложнения мозга в корково-подкорковых отношениях корково-таламические связи становятся определяющими, а в них, в свою очередь, преобладающими (по сравнению с проекционными, релейными) становятся ассоциативные связи. Так, у обезьян проекционные зоны занимают еще значительно большую территорию коры головного мозга по сравнению с ассоциативными зонами; напр., только затылочная проекционная зона составляет ок. 20% площади всей коры. У человека соотношения обратные — доминирующими являются ассоциативные зоны, и затылочная проекционная зона составляет всего 12% площади новой коры. Поэтому и в таламических комплексах мозга человека наиболее развиты ассоциативные ядра.
Специфические ядра
Специфические ядра зрительного бугра имеют ряд отличительных особенностей. Все образования этой группы получают сенсорную информацию от вторых нейронов (нервных клеток) чувствительных проводящих путей. Второй нейрон, в свою очередь, может быть расположен в спинном мозге или в одной из структур ствола мозга: продолговатом мозге, мосту, среднем мозге.
Каждый из сигналов, поступающих снизу, обрабатывается в таламусе и далее идет в соответствующую область коры. В какую именно область поступает нервный импульс, зависит от того, какую информацию он несет. Так, информация о звуках поступает в слуховую кору, об увиденных предметах — в зрительную и так далее.
Помимо импульсов из вторых нейронов проводящих путей, специфические ядра отвечают за восприятие информации, поступающей из коры, ретикулярной формации, ядер ствола мозга.
Ядра, которые находятся в передней части таламуса, обеспечивают проведение импульсов из лимбической коры головного мозга через гиппокамп и гипоталамус. После обработки информации она опять поступает в лимбическую кору. Таким образом, нервный импульс циркулирует по определенному кругу.
Физиология
Исследование структуры и функции таламических образований с помощью современных комплексных методов выявило чрезвычайно сложный характер отношений Т. как с выше расположенными отделами переднего мозга, так и с находящимися ниже структурами мозгового ствола, мозжечка и спинного мозга. Тесная интеграция соматических, висцеральных, мотивационных влияний, осуществляемая на уровне таламических комплексов, определяет ведущую роль этих образований промежуточного мозга в формировании не только корковоподкорковых отношений, но и в становлении и развитии целостных поведенческих актов животных и человека.
Изучение функций Т. проводят путем его электрического раздражения или разрушения в сочетании с анализом динамики различных поведенческих реакций, регистрацией ЭЭГ, вызванных потенциалов или электрической активности одиночных нейронов в ответ на различные периферические раздражения в экспериментах на животных и в леч. целях — у человека.
Установлено, что в релейных ядрах Т. переключаются импульсы тактильной, кинестетической, температурной, болевой (локализованная боль), вкусовой и висцеральной чувствительности (см.). В каждое ядро поступают импульсы с противоположной стороны тела, лишь область лица имеет в заднем вентральном ядре Т. билатеральное представительство. Части тела, выполняющие наиболее интенсивные сенсорно-двигательные функции (лицо, язык, дистальные отделы конечностей), имеют более обширное представительство. В медиальный отдел заднего вентрального ядра, соматотопическая организация к-рого хорошо изучена, приходит афферентация от ростральных частей тела и импульсы от вкусовых рецепторов, в латеральный отдел — от рецепторов туловища и конечностей. Проекции тактильной и висцеральной чувствительности имеют в Т. широкое перекрытие, что может обусловливать феномен иррадиации висцеральной боли (см.) на поверхности тела. Общие проекции в Т. имеют также тактильная и глубокая (проприоцептивная) чувствительность. Тонкодифференцированная афферентная импульсация от термо- и ноцирецепторов проходит преимущественно по спиноталамическому пути в заднее вентральное ядро, тогда как более грубые формы этих видов чувствительности (диффузная боль, дифференцировка больших температурных диапазонов) связаны преимущественно с неспецифическими интраламинарными ядрами Т. (парафасцикулярным комплексом), а также, возможно, с задними ядрами. Электростимуляция заднего вентрального ядра у человека в процессе стереотаксических операций (см. Стереотаксическая нейрохирургия, Стереотаксический метод) вызывает парестезии, реже нарушение схемы тела (см. Схема тела).
К несенсорным релейным образованиям относят переднюю группу ядер, передающую импульсацию от сосцевидных тел к лимбической области коры, а также два ядра вентральной группы Т. — переднее вентральное и латеральное вентральное, передающие основную массу возбуждений из мозжечка и бледного шара к моторным отделам коры головного мозга, что и определяет их роль в регуляции непроизвольной двигательной активности и мышечного тонуса. Указанные связи латерального вентрального ядра обосновывают целесообразность его деструкции у больных паркинсонизмом и с различными гиперкинезами.
Группа ассоциативных ядер Т. имеет основным источником афферентации релейные и отчасти неспецифические структуры Т. и гипоталамус. В последнее время показано, что в этих ядрах может заканчиваться часть сенсорных проекций (соматосенсорных, слуховых, зрительных, висцеральных). Взаимодействие различных афферентных импульсов на нейронах ассоциативных ядер Т., их сложные внутриталамические и межталамические связи объясняют важную роль этой группы образований в интегративной деятельности мозга. Различают таламопариетальную и таламофронтальную ассоциативные системы. Первая имеет преимущественное отношение к формированию комплексов афферентных воздействий, необходимых для оценки сигнальной значимости сложных видов раздражителей, лежащей в основе гностической функции, особенно зрительного и слухового восприятия, а также сенсорного обеспечения управления произвольными движениями. Медиальное дорсальное ядро принадлежит таламофронтальной ассоциативной системе, участвующей прежде всего в аффективно-эмоциональной и программирующей деятельности. Стимуляция медиального дорсального ядра Т. у больных вызывает разнообразные положительные и отрицательные эмоциональные реакции; деструкция этого ядра в леч. целях, а также в экспериментах на животных ослабляет чувство страха, тревоги и напряженности, но при этом возникают нерезко выраженные симптомы так наз. лобного синдрома — снижение инициативы и интеллектуальных способностей на фоне стойкого уменьшения эмоциональной реактивности. При распространении патол. процесса на медиальные отделы Т. наблюдают развитие слабоумия (см.).
Группа неспецифических ядер Т. получает афферентацию прежде всего из ретикулярных образований мозгового ствола, однако есть данные о возможности окончания в них волокон, идущих в составе специфических восходящих систем. Хотя ядра этой группы проецируются на кору более диффузно, чем первые две группы, нек-рым из них свойствен более или менее локальный характер связей с корой и полосатым телом (стриопаллидумом). К этой группе относят внутрипластинчатые (интраламинарные) ядра (парафасцикулярное ядро, латеральное центральное и парацентральное ядра), ретикулярное ядро таламуса, ядра средней линии Т. Открытие неспеци-фицеских ядер Т. связано с работами Демпси и Морисон (E. W. Dempsey, R. S. Morison, 1943), показавшими, что при раздражении этих ядер в коре головного мозга возникает распространенная реакция вовлечения. Позднее С. П. Нарикашвили, Э. С. Мониава, В. И. Гусельников и А. Я. Супин (1968) и др. обнаружили, что в генезе этой реакции, так же как и реакции усиления, ведущую роль играет взаимодействие несиецифических ядер Т. со специфическими его образованиями.
Данные о функциональном значении ядер этой группы разноречивы: с одной стороны, установлено, что разрушение комплекса парафасцикулярное ядро — центральное срединное ядро (срединный центр) влияет на позитивное и негативное обучение крыс; с другой стороны, эффект повреждения этого комплекса в значительной степени зависит от предшествующего эмоционального фона; показана также важная роль афферентации данного комплекса в деятельности нейронов хвостатого ядра (см. Базальные ядра). Представление о неспецифичности этой группы ядер относительно. На основании данных об их строении, связях и функции можно говорить о существовании в Т. сложных комплексов, образованных релейными и неспецифическими ядрами. Изучение функций ядер средней линии затруднено их теснейшей близостью к ассоциативным и релейным ядрам Т. Разрушение медиальных отделов Т. может сопровождаться нарушениями условных рефлексов (см.), памяти (см.), сна (см.).
В группе неспецифических ядер особое место занимает ретикулярное ядро Т. Последнее примыкает снаружи к различным релейным ядрам дорсального Т. и частично имеет общую с ними проекцию на кору. Предполагают, что это ядро является посредником как в деятельности других ядер Т., так и в таламо-кортикальном взаимодействии его специфических и неспецифических образований. В наст, время очевидна важная роль Т. в осуществлении сложных функций восприятия и переработки различных сигналов, поступающих далее в кору головного мозга и базальные ядра, в поддержании уровня бодрствования и эмоционального состояния, в нормальном обеспечении простых и более сложных форм поведения и памяти. Полное двустороннее удаление Т. у собак и кошек обнаруживает резкий дефицит приспособительных реакций, нарушения чувствительности и чередований цикла бодрствование — сон. Оперативная изоляция коры и базальных ядер от Т. и мозгового ствола (так наз. таламическое животное) приводит к массивной ретроградной дегенерации нейронов Т. и сопровождается резкими и необратимыми изменениями адаптивного поведения, вплоть до невозможности самостоятельного питания.
В последнее время в связи с накоплением данных о межполушарной асимметрии появились указания на функциональную неравнозначность правого и левого Т. у человека. Так, использование вербализуемого материала в тестах на кратковременную память, видимо, осуществляется с участием преимущественно левого Т., невербализуемого материала — с участием правого Т.; наиболее существенным синдромом левостороннего поражения таламуса является несодержательность речи.
В 70-х гг. был сформулирован ряд новых принципов таламокортикального взаимодействия, имеющих отношение к пониманию интеграции нервных процессов на уровне переднего мозга: принципы «ядерного» и «рассеянного» типов таламокортикальных проекций, переслаивания этих различных проекций между собой, их различного распределения в проекционных и ассоциативных областях новой коры, моно- и оли-госинаптической передачи возбуждений из Т. в кору, а также топографического соответствия (преимущественно для релейных и ассоциативных структур) и несоответствия (гл. обр. для неспецифических структур Т.) таламокортикальных и кортико-таламических влияний. В нейрофизиол. исследованиях обнаружена динамическая интеграция специфических и неспецнфических влияний на диэнцефальном и корковом уровнях, установлено, что ритмическая активность коры головного мозга обусловлена активностью таламических ядер, что формирование a-ритма в коре происходит при участии как таламических, так и корковых структур. Обнаружено, что нейрофизиол. основой ритмической активности головного мозга являются механизмы возвратного торможения (см.) п возбуждения (см.), разыгрывающиеся на таламическом и корковом уровнях. Выделены и изучены новые пути взаимоотношений ассоциативных и проекционных таламокортикальных систем и сформулировано положение о сложно функционирующих таламокортико-таламических ассоциативных системах. Получила новое освещение проблема корковой регуляции деятельности таламических ядер: показано, что регуляция релейных таламических ядер осуществляется гл. обр. прямыми кортико-таламическими путями, а неспецифических ядер — преимущественно с участием ретикулярной формации мозгового ствола. Установлено, что ретикулярные системы мозга способны дифференцировать афферентные сигналы и направлять их в соответствующие структуры мозга под влиянием кортико-фугальной модуляции.
Ассоциативные ядра
Ассоциативные ядра находятся ближе к задне-медиальной части таламуса, а также в области подушки. Особенность этих структур в том, что они не участвуют в восприятии информации, которая подходит из нижележащих образований центральной нервной системы. Эти ядра необходимы для получения уже обработанных сигналов в других ядрах таламуса или в вышележащих мозговых структурах.
Суть «ассоциативности» этих ядер в том, что к ним подходят любые сигналы, а нейроны способны их адекватно воспринять. Сигналы из этих структур поступают в области коры с соответствующим названием — ассоциативные зоны. Они расположены в височной, лобной и теменной частях коры. Благодаря поступлению этих сигналов человек способен:
- узнавать предметы;
- связывать речь с движениями и увиденными предметами;
- осознавать положение своего тела в пространстве;
- воспринимать пространство трехмерным и прочее.
Нейроны
Они проявляют высокую чувствительность к фармакологическим средствам и гормонам. Такие препараты, как «Резерпин», «Аминазин», «Серпазил» и прочие способны снизить активность формации. В нейронах происходит взаимодействие восходящих и нисходящих сигналов. Импульсы находятся в постоянной циркуляции в цепях. За счет этого поддерживается активность. Она, в свою очередь, необходима для поддержания тонуса нервной системы. В случае разрушения формации, в особенности верхних ее участков, наступает глубокий сон, хотя афферентные сигналы продолжают поступать в кору по другим путям.
Неспецифические ядра
Неспецифическими эта группа ядер называется потому, что получает информацию практически от всех структур центральной нервной системы:
- ретикулярной формации;
- ядер экстрапирамидной системы;
- других ядер зрительного бугра;
- стволовых структур мозга;
- образований лимбической системы.
Импульс от неспецифических ядер также идет ко всем областям коры головного мозга. Такая выборочность, как в случае с ассоциативными и специфическими ядрами, здесь отсутствует.
Так как именно эта группа ядер имеет наибольшее количество связей, считается, что благодаря ей обеспечивается слаженная, координированная работа всех участков головного мозга.
Отдельно о зрительном бугре
Ранее считалось, что таламус обрабатывает только зрительные импульсы, тогда орган и получил название — зрительные бугры. Сейчас это название считается устаревшим, так как орган обрабатывает практически весь спектр афферентных систем (кроме обоняния).
Система, которая обеспечивает зрительное восприятие – одна из самых интересных. Основной внешний орган зрения – глаз – рецептор, который имеет сетчатку и оснащен особенными клетками (колбочки, палочки), которые трансформируют световой пучок и электрический сигнал. Электрический сигнал, в свою очередь, проходя по нервным клеткам, попадает в латеральный центр таламуса, который отправляет обработанный сигнал в центральный отдел коры головного мозга. Тут происходит окончательный анализ сигнала, благодаря чему формируется увиденное, то есть – картинка.
Метаталамус
Отдельно выделяют группу ядер зрительного бугра под названием метаталамус. Данная структура состоит из медиального и латерального коленчатых тел.
Медиальное коленчатое тело получает информацию о слухе. Из нижележащих отделов мозга информация поступает через верхние горбики среднего мозга, а сверху структура получает импульс из слуховой области коры.
Латеральное коленчатое тело относится к зрительной системе. Чувствительная информация к ядрам этой группы поступает от сетчатки глаза через зрительные нервы и зрительный тракт. Обработанная в таламусе информация далее идет к затылочной области коры, где находится первичный центр зрения.
Функции таламуса
Как происходит обработка поступающей с периферии чувствительной информации, которая далее передаются в кору переднего мозга? Это и есть основная роль зрительного бугра.
Благодаря этой функции при повреждении коры возможно восстановление чувствительности посредством таламуса. Таким образом, возможна репарация болевого, температурного чувства, а также грубого осязания.
Еще одна важная функция таламуса — это координация движений и чувствительности, то есть сенсорной и моторной информации. Это обусловлено тем, что в таламус поступают не только сенсорные импульсы. Также к нему идут импульсы от мозжечка, ганглиев экстрапирамидной системы, коры большого мозга. А эти структуры, как известно, принимают участие в осуществлении движений.
Также зрительный бугор участвует в поддержании сознательной активности, регуляции сна и бодрствования. Эта функция осуществляется за счет наличия связей с голубым пятном ствола мозга и гипоталамусом.
Гипоталамус мозга
Строение гипоталамуса очень сложное, поэтому в этой статье будут рассмотрены только его функции. Они заключаются в поведенческих ответных реакциях человека, а также во влиянии на вегетатику. Помимо этого, гипоталамус активно принимает участие в регенерации резервов.
Гипоталамус тоже имеет множество ядер, которые делятся на задние, средние и передние. Ядра задней категории регулируют симпатические реакции организма – повышение давление, учащенный пульс, расширение зрачка глаза. Ядра средней категории наоборот, снижают симпатические проявления.
Гипоталамус отвечает за:
- терморегуляцию;
- чувство насыщения и голода;
- страх;
- половое влечение и так далее.
Все эти процессы происходят в результате активации или торможения различных ядер.
К примеру, если у человека расширяются сосуды, и ему становится холодно, значит произошло раздражение передней группы ядер, а если повреждаются ядра заднего порядка, то это может спровоцировать летаргический сон.
Гипоталамус отвечает за регуляцию движений, если в этой области происходит возбуждение, человек может совершать хаотические движения. Если нарушения происходят в так называемом сером бугре, который тоже входит в состав гипоталамуса, то человек начинает страдать от нарушения обменных процессов.
Симптомы поражения
Так как через таламус проходят практически все сигналы от других структур нервной системы, поражение зрительного бугра может проявляться массой симптомов. Обширное поражение таламуса можно диагностировать по следующим клиническим признакам:
- нарушение чувствительности, в первую очередь — глубокой;
- жгучие, резкие боли, которые сначала появляются при прикосновении, а потом и спонтанно;
- нарушения моторики, среди которых встречается так называемая таламическая кисть, проявляющаяся чрезмерным сгибанием пальцев в пястно-фаланговых и разгибанием в межфаланговых суставах;
- зрительные расстройства — гемианопсия (выпадение полей зрения с противоположной от поражения стороны).
Таким образом, таламус — важная структура головного мозга, которая обеспечивает интеграцию всех процессов в организме.
Патологии гипоталамуса
Все недуги гипоталамуса связаны с нарушением функции этой структуры, а точнее с особенностями гормонального синтеза. Заболевания могут возникнуть по причине избыточного продуцирования гормонов, по причине сниженной секреции гормонов, но также недуги могут появиться при нормальной выработке гормонов гипоталамуса. Существует очень тесная связь между гипоталамусом и гипофизом – у них общее кровообращение, похожее анатомическое строение и идентичные функции. Поэтому часто заболевания объединяют в одну группу, которую именуют патологиями гипоталамо-гипофизарной системы.
Нередко причиной возникновения патологической симптоматики является возникновение аденомы гипофиза или самого гипоталамуса. В этом случае гипоталамус начинает продуцировать большое количество гормонов, в результате чего и появляется соответствующая симптоматика.
Типичным поражение гипоталамуса являет пролактинома – опухоль, которая является гормонально активной, так как вырабатывает пролактин.
Еще одним опасным заболеванием является гипоталамо-гипофизарный синдром, этот недуг связан с нарушением функциональности как гипофиза, так и гипоталамуса, что приводит к развитию характерной клинической картины.
В связи с тем, что заболеваний, поражающих гипоталамо-гипофизарную систему много, ниже будут приведены общие симптомы, по которым можно заподозрить патологии этого отдела головного мозга:
- Проблемы с насыщением организма. Ситуация может развиваться в двух направлениях – либо человек полностью теряет аппетит, либо не чувствует насыщения сколько бы не съел.
- Проблемы с терморегуляцией. Проявляется это в повышении температуры, при этом никаких воспалительных процессов в организме не наблюдается. Кроме того, повышение температурных показателей сопровождается ознобом, повышенным потоотделением, повышенной жаждой, ожирением и неконтролируемым голодом.
- Эпилепсия по гипоталамическому признаку – перебои в работе сердца, повышенное артериальное давление, болевые ощущения в эпигастральной области. При приступе человек теряет сознание.
- Изменения в работе вегетососудистой системе. Они проявляются в работе пищеварения (отрыжка, боли в животе, срывы стула), в работе системы дыхания (тахипноэ, затрудненный вдох, удушье) и в работе сердца и сосудов (сбои в сердечном ритме, высокое или низкое артериальное давление, загрудинные боли).
Лечением заболеваний гипоталамуса занимаются неврологи, эндокринологи и гинекологи.