Neurons of the cerebral cortex
Cerebral cortex
or
cerebral cortex
(lat. cortex cerebri) - a structure of the brain, a layer of gray matter 1.3-4.5 mm thick[1], located along the periphery of the cerebral hemispheres and covering them. The greatest thickness is observed in the upper parts of the precentral, postcentral gyri and paracentral lobule[2].
The cerebral cortex plays a very important role in the implementation of higher nervous (mental) activity [2].
In humans, the cortex makes up on average 44% of the volume of the entire hemisphere as a whole [2]. The surface area of the cortex of one hemisphere in an adult is on average 2200 cm²[2]. The superficial parts account for 1/3, and those lying deep between the gyri account for 2/3 of the total area of the cortex [1].
The size and shape of the grooves are subject to significant individual fluctuations - not only the brains of different people, but even the hemispheres of the same individual are not quite similar in the pattern of the grooves[1].
The entire cerebral cortex is usually divided into 4 types: ancient (paleocortex), old (archicortex), new (neocortex) and intermediate cortex (consisting of intermediate ancient and intermediate old cortex). The surface of the neocortex in humans occupies 95.6%, the archicortex - 2.2%, the paleocortex - 0.6%, the intermediate - 1.6%[2].
Content
- 1 Anatomy 1.1 Frontal lobe
- 1.2 Parietal lobe
- 1.3 Temporal lobe
- 1.4 Insula (islet)
- 1.5 Grooves and convolutions of the medial surface
- 1.6 Grooves and convolutions of the lower surface
- 2.1 Structure
Midbrain
The midbrain is located in the stem part. It is a conductor of signals from the front to various departments. Its main function is to regulate muscle tone. It is also responsible for the transmission of tactile sensations, coordination and reflexes. The functions of parts of the human brain depend on their location. For this reason, the midbrain is responsible for the vestibular apparatus. Thanks to the midbrain, a person can simultaneously perform several functions.
In the absence of intellectual activity, brain function is disrupted. People over 70 years of age are susceptible to this. When the middle part malfunctions, coordination failures occur and visual and auditory perception shifts.
Anatomy
Lobes of the cerebral hemispheres
The cerebral cortex covers the surface of the hemispheres and forms a large number of grooves of varying depth and length (lat. sulci cerebri). Between the grooves there are cerebral gyri of varying sizes (lat. gyri cerebri) [3].
In each hemisphere the following surfaces are distinguished:
- convex superolateral surface
(lat. facies superolateralis), adjacent to the inner surface of the bones of the cranial vault - the lower surface
(lat. facies inferior), the anterior and middle sections of which are located on the inner surface of the base of the skull, in the area of the anterior and middle cranial fossae, and the posterior ones - on the tentorium of the cerebellum - the medial surface
(lat. facies medialis), directed towards the longitudinal fissure of the brain [3].
These three surfaces of each hemisphere, passing one into another, form three edges. The superior edge (lat. margo superior) separates the superolateral and medial surfaces. The inferolateral edge (lat. margo inferolateralis) separates the superolateral surface from the bottom. The inferomedial edge (lat. margo inferomedialis) is located between the lower and medial surfaces [3].
In each hemisphere, the most prominent places are distinguished: in front - the frontal pole (lat. polus frontalis), in the back - the occipital pole (lat. polus occipitalis), and on the side - the temporal pole (lat. polus temporalis) [3].
The hemisphere is divided into five lobes. Four of them are adjacent to the corresponding bones of the cranial vault:
- frontal lobe (lat. lobus frontalis)
- parietal lobe (lat. lobus parietalis)
- occipital lobe (lat. lobus occipitalis)
- temporal lobe (lat. lobus temporalis)
- insular lobe (lat. lobus insularis) (islet) (lat. insula) - is located deep in the lateral fossa of the cerebrum (lat. fossa lateralis cerebri), separating the frontal lobe from the temporal lobe [3].
Frontal lobe
Frontal lobe
The frontal lobe is separated from the parietal lobe by a deep central (Rolandic) groove (lat. sulcus centralis). It begins on the medial surface of the hemisphere, passes to its superolateral surface, runs along it slightly obliquely, from back to front, and usually does not reach the lateral (lateral or Sylvian) fissure of the brain [3].
Approximately parallel to the central sulcus is the precentral sulcus (lat. sulcus precentralis), which does not reach the upper edge of the hemisphere. The precentral sulcus borders the front of the precentral gyrus (lat. gyrus precentralis) [3].
The superior and inferior frontal sulci (lat. sulci frontales superior et inferior) are directed forward from the precentral sulcus. They divide the frontal lobe into:
- superior frontal gyrus (lat. gyrus frontalis superior), which is located above the superior frontal sulcus and passes to the medial surface of the hemisphere
- the middle frontal gyrus (lat. gyrus frontalis medius), which is bounded by the superior and inferior frontal sulci. The orbital (anterior) segment of this gyrus passes onto the inferior surface of the frontal lobe
- the inferior frontal gyrus (lat. gyrus frontalis inferior), which lies between the inferior frontal sulcus and the lateral sulcus of the brain and the branches of the lateral sulcus is divided into a number of parts [3]
The lateral sulcus (lat. sulcus lateralis) is one of the deepest grooves in the brain. It separates the temporal lobe from the frontal and parietal lobes. The lateral groove lies on the superolateral surface of each hemisphere and runs from top to bottom and anteriorly. In the depths of this groove there is a depression - the lateral fossa of the cerebrum (lat. fossa lateralis cerebri), the bottom of which is the outer surface of the insula [3].
From the lateral sulcus, small grooves called rami extend superiorly. The most constant of them are the ascending (lat. ramus ascendens) and anterior (lat. ramus anterior) branches. The superior posterior section of the groove is called the posterior branch (lat. ramus posterior)[3].
The inferior frontal gyrus, within which the ascending and anterior branches pass, is divided into three parts:
- posterior - tegmental part (lat. pars opercularis), limited in front by the ascending branch
- middle - triangular part (lat. pars triangularis), lying between the ascending and anterior branches
- anterior - orbital part (lat. pars orbitalis), located between the anterior branch and the inferolateral edge of the frontal lobe[3]
Parietal lobe
The parietal lobe
lies posterior to the central sulcus, which separates it from the frontal. It is delimited from the temporal sulcus by the lateral sulcus of the brain, from the occipital sulcus by part of the parieto-occipital sulcus (lat. sulcus parietooccipitalis)[3].
The postcentral gyrus (lat. gyrus postcentralis) runs parallel to the precentral gyrus. From it posteriorly, almost parallel to the longitudinal fissure of the cerebrum, there is an intraparietal groove (lat. sulcus intraparietalis), dividing the postero-superior parts of the parietal parts of the parietal lobe into two gyri: the upper (lat. lobulus parietalis superior) and the lower (lat. lobulus parietalis inferior) parietal lobules. In the inferior parietal lobe, two relatively small gyri are distinguished: the supramarginal gyrus (lat. gyrus supramarginalis), which lies in front and closes the posterior sections of the lateral sulcus, and the angular gyrus (lat. gyrus angularis), located posterior to the previous one, which closes the superior temporal sulcus [3].
Between the ascending and posterior branches of the lateral sulcus of the brain there is a section of the cortex designated as the frontoparietal operculum (lat. operculum frontoparietalis). It includes the posterior part of the inferior frontal gyrus, the lower parts of the precentral and postcentral gyri, as well as the lower part of the anterior part of the parietal lobe [3].
Temporal lobe
The temporal lobe
has the most pronounced boundaries. It has a convex lateral surface and a concave bottom surface. The obtuse pole of the temporal lobe faces forward and slightly downward. The lateral cerebral sulcus sharply demarcates the temporal lobe from the frontal lobe[3].
Two grooves located on the superolateral surface: the upper (lat. sulcus temporalis superior) and the lower (lat. sulcus temporalis inferior) temporal sulci, following almost parallel to the lateral sulcus of the brain, divide the lobe into three temporal gyri: upper, middle and lower (lat. gyri temporales superior, medius et inferior)[3].
Those parts of the temporal lobe that are directed towards the lateral groove of the brain are cut by short transverse temporal grooves (lat. sulci temporales transversi). Between these grooves lie 2-3 short transverse temporal gyri, associated with the gyri of the temporal lobe (lat. gyri temporales transversi) and the insula [3].
Insula (islet)
It lies at the bottom of the lateral fossa of the cerebrum (lat. fossa lateralis cerebri).
It is a three-sided pyramid, facing its apex - the pole of the insula - anteriorly and outwardly, towards the lateral sulcus. From the periphery, the insula is surrounded by the frontal, parietal and temporal lobes, which participate in the formation of the walls of the lateral sulcus of the brain [3].
The base of the islet is surrounded on three sides by a circular groove of the islet (lat. sulcus circularis insulae).
Its surface is cut by a deep central groove of the insula (lat. sulcus centralis insulae). This groove divides the insula into anterior and posterior parts[3].
On the surface there are a large number of small convolutions of the insula (lat. gyri insulae). The large anterior part consists of several short convolutions of the insula (lat. gyri breves insulae), the posterior part consists of one long convolution (lat. gyrus longus insulae)[3].
Furrows and convolutions of the medial surface
The frontal, parietal and occipital lobes extend onto the medial surface of the hemisphere.
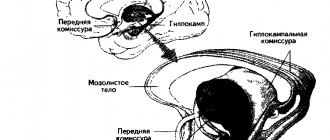
The cingulate gyrus (lat. gyrus cinguli) begins with the subcallosal area (lat. area subcallosa), goes around the corpus callosum and, through a narrow strip - the isthmus of the cingulate gyrus (lat. isthmus gyri cinguli) passes into the parahippocampal gyrus on the lower surface of the hemisphere [3].
The sulcus of the corpus callosum (lat. g sulcus corporis callosi) separates the cingulate gyrus from the corpus callosum and on the lower surface of the hemisphere continues into the sulcus of the hippocampus [3].
The cingulate gyrus is bounded above by the cingulate groove (lat. sulcus cinguli). In the latter, there is a front part convex towards the frontal pole and a back part, which, following along the cingulate gyrus and not reaching its posterior section, rises to the upper edge of the cerebral hemisphere. The posterior end of the sulcus lies behind the upper end of the central sulcus. Between the precentral sulcus, the end of which is sometimes clearly visible at the upper edge of the medial surface of the hemisphere, and the end of the cingulate sulcus, there is a paracentral lobule (lat. lobulus paracentralis)[3].
Above the cingulate gyrus, in front of the subcallosal field, the medial frontal gyrus (lat. gyrus frontalis medialis) begins. It extends to the paracentral lobule and is the lower part of the superior frontal gyrus.
Behind the cingulate groove lies a small quadrangular lobe - the precuneus (lat. precuneus). Its posterior border is the deep parieto-occipital sulcus (lat. sulcus parietooccipitalis), the lower border is the subparietal sulcus (lat. sulcus subparietalis), separating the precuneus from the posterior part of the cingulate gyrus [3].
Behind and below the precuneus lies a triangular lobe - wedge (lat. cuneus). The convex outer surface of the wedge participates in the formation of the occipital pole. The apex of the wedge, directed downward and forward, almost reaches the posterior part of the cingulate gyrus. The posteroinferior border of the wedge is the very deep calcarine groove (lat. sulcus calcarinus), the anterior border is the parieto-occipital groove [3].
Grooves and convolutions of the lower surface
On the lower surface of the frontal lobe there is an olfactory groove (lat. sulcus olfactorius). Inwardly from it, between it and the inferomedial edge of the hemisphere, lies the straight gyrus (lat. gyrus rectus). Its posterior section reaches the anterior perforated substance (lat. substantia perforata anterior). Outside the groove is the rest of the lower surface of the frontal lobe, cut by short orbital grooves (lat. sulci orbitales), into a number of small orbital gyri (lat. gyri orbitales)[3].
The lower surface of the temporal lobe is separated from the cerebral peduncles by the deep groove of the hippocampus (lat. sulcus hippocampi). In the depths of the furrow lies a narrow serrated gyrus (lat. gyrus dentatus). Its anterior end passes into the hook, and the posterior end into the ribbon gyrus (lat. gyrus fasciolaris) lying under the splenium of the corpus callosum. Lateral to the sulcus is the parahippocampal gyrus (lat. gyrus parahippocampalis). Anteriorly, this gyrus has a thickening in the form of a hook (lat. uncus), and posteriorly it continues into the lingual gyrus (lat. gyrus lingualis). The parahippocampal and lingual gyri are limited on the lateral side by the collateral groove (lat. sulcus collateralis), which passes in front into the nasal groove (lat. sulcus rhinalis). The rest of the lower surface of the temporal lobe is occupied by the medial and lateral occipitotemporal gyri (lat. gyri occipitotemporales medialis et lateralis), separated by the occipitotemporal sulcus (lat. sulcus occipitotemporalis). The lateral occipitotemporal gyrus is separated by the inferolateral edge of the hemisphere from the inferior temporal gyrus [3].
Functions of the hemispheres
The right and left hemispheres of humans perform different functions. The left hemisphere contains the centers of oral and written language. Here the processes of analysis and synthesis of information are carried out, generalizations are made and decisions are made. Verbal and logical thinking provided by the left hemisphere allows one to cognize the essence of an object and go beyond the limits of the individual world. On its basis human knowledge is formed. It is passed down from generation to generation by recording verbal or symbolic signals.
The right hemisphere carries out imaginative thinking. Operating with images of objects in the external world, it can create unprecedented, fantastic combinations from them. And this is the basis of creativity and making unusual decisions. The right hemisphere is extremely important for musical and artistic creativity. It is known that the most outstanding artists, poets, and musicians are people with a predominance of right-hemisphere thinking.
Despite the functional asymmetry, the brain works as a single whole. By summarizing information, it ensures adequate behavior, thinking, consciousness, memory, labor and creative activity of a person.
Histology
Structure
Cytoarchitecture (cell arrangement)
- molecular layer
- outer granular layer
- layer of pyramidal neurons
- inner granular layer
- ganglion layer (inner pyramidal layer; Betz cells)
- layer of polymorphic cells
Myeloarchitecture (arrangement of fibers)
- molecular layer strip
- strip of outer granular layer
- strip of inner granular layer
- strip of the ganglion layer [4].
The cerebral cortex is represented by a layer of gray matter with an average thickness of about 3 mm (1.3 - 4.5 mm). It is most strongly developed in the anterior central gyrus. The abundance of furrows and convolutions significantly increases the area of gray matter in the brain. The cortex contains about 10-14 billion nerve cells. Its various sections, which differ from each other in certain features of the location and structure of cells (cytoarchitectonics), the arrangement of fibers (myeloarchitectonics) and functional significance, are called fields. They represent places of higher analysis and synthesis of nerve impulses. There are no sharply defined boundaries between them. The cortex is characterized by an arrangement of cells and fibers in layers [5].
Typical of the new cortex (lat. neocortex) is the presence of six layers, differing from each other mainly in the shape of the nerve cells included in them. At the same time, on the medial and lower surfaces of the hemispheres, sections of the old (lat. archipallium) and ancient (lat. paleopallium) cortex, which has a 2-layer and 3-layer structure, have been preserved [1]. An intermediate bark (lat. mesopallium) is also distinguished, located between the old and new, as well as ancient and new bark [3]. The ancient cortex is represented by the hippocampus, and the old cortex is represented by a section of the cortex near the olfactory bulb on the lower surface of the frontal lobe[1].
Cytoarchitecture
Multipolar neurons of the cerebral cortex are very diverse in shape. Among them are:
- pyramidal
- stellate
- fusiform
- arachnids
- horizontal
Pyramidal neurons constitute the main and most specific form for the cerebral cortex (80-90% of all neurons). Their sizes vary from 10 to 140 microns. They have an elongated triangular body, the apex of which faces the surface of the bark. Dendrites extend from the apex and lateral surfaces of the body, ending in various layers of gray matter. Axons originate from the base of pyramidal cells; in some cells they are short, forming branches within a given area of the cortex, in others they are long, entering the white matter [5].
Pyramidal cells of different layers of the cortex differ in size and have different functional significance. Small cells are interneurons, the axons of which connect individual areas of the cortex of one hemisphere (associative neurons) or two hemispheres (commissural neurons). These cells are found in varying numbers in all layers of the cortex. The human cerebral cortex is especially rich in them. Axons of large pyramidal neurons take part in the formation of pyramidal tracts that project impulses to the corresponding centers of the brain stem and spinal cord [5].
The neurons of the cortex are located in vaguely delimited layers. Each layer is characterized by the predominance of one type of cell. There are 6 main layers in the motor cortex:
- Molecular (lat. lamina molecularis)
- External granular (lat. lamina granularis externa)
- Pyramidal neurons (lat. lamina pyramidalis)
- Internal granular (lat. lamina granularis interna)
- Ganglionic (layer of Betz cells) (lat. lamina ganglionaris)
- Layer of multiform (polymorphic) cells (lat. lamina multiformis) [5]
The cerebral cortex also contains a powerful neuroglial apparatus that performs trophic, protective, supporting and delimiting functions [5].
On the medial and lower surface of the hemispheres, sections of the old, ancient cortex have been preserved, which have a two-layer and three-layer structure.
Molecular layer
The molecular layer of the cortex contains a small number of small spindle-shaped association cells. Their axons run parallel to the surface of the brain as part of the tangential plexus of nerve fibers of the molecular layer. The bulk of the fibers of this plexus are represented by the branching of dendrites of neurons in the underlying layers [5].
Outer granular layer
The outer granular layer is formed by small neurons with a diameter of about 10 microns, having a round, angular and pyramidal shape, and stellate neurons. The dendrites of these cells rise into the molecular layer. Axons either go into the white matter or, forming arcs, also enter the tangential plexus of fibers of the molecular layer [5].
Layer of pyramidal neurons
It is the widest compared to other layers of the cerebral cortex. It is especially well developed in the precentral gyrus. The size of pyramidal cells consistently increases within 10-40 µm from the outer zone of this layer to the inner one. The main dendrite extends from the top of the pyramidal cell and is located in the molecular layer. The dendrites, originating from the lateral surfaces of the pyramid and its base, are of insignificant length and form synapses with adjacent cells of this layer. The axon of a pyramidal cell always extends from its base. In small cells it remains within the cortex; an axon belonging to a large pyramid usually forms a myelin association or commissural fiber that goes into the white matter [5].
Inner granular layer
In some fields of the cortex it is very developed (for example, in the visual cortex). However, in other areas it may be absent (in the precentral gyrus). This layer is formed by small stellate neurons. It contains a large number of horizontal fibers [5].
Ganglion layer (Inner pyramidal layer; Betz cells)
It is formed by large pyramidal cells, and the area of the precentral gyrus contains giant cells, first described by the Russian anatomist V. A. Betz in 1874 (Betz cells). They reach a height of 120 and a width of 80 microns. Unlike other pyramidal cells of the cortex, giant Betz cells are characterized by the presence of large clumps of chromatophilic substance. Their axons form the main part of the corticospinal and corticonuclear tracts and end on motor neurons of the brain stem and spinal cord [5].
Before leaving the cortex, many collaterals depart from the pyramidal tract. Axons from Betz giant cells give rise to collaterals that send inhibitory impulses to the cortex itself. Also, collaterals of fibers of the pyramidal tract go to the striatum, red nucleus, reticular formation, pontine and inferior olivary nuclei. The pontine and inferior olivary nuclei transmit signals to the cerebellum. Thus, when the pyramidal tract transmits a signal that causes goal-directed movement to the spinal cord, the basal ganglia, brain stem, and cerebellum simultaneously receive signals. In addition to the collaterals of the pyramidal tracts, there are fibers that go directly from the cortex to the intermediate nuclei: the caudate body, the red nucleus, the nuclei of the reticular formation of the brain stem, etc.[5]
Multimorph cell layer
Formed by neurons of various, predominantly spindle-shaped shapes. The outer zone of this layer contains larger cells. The neurons of the inner zone are smaller and lie at a greater distance from each other. The axons of the polymorphic layer cells extend into the white matter as part of the efferent pathways of the brain. Dendrites reach the molecular layer of the cortex [5].
Myeloarchitecture
Among the nerve fibers of the cerebral cortex we can distinguish:
- association fibers - connect individual areas of the cortex of one hemisphere
- commissural fibers - connect the cortex of the two hemispheres
- projection fibers - connect the cortex with the nuclei of the lower parts of the central nervous system. Afferent projection fibers end in the layer of pyramidal neurons[5]
In addition to the tangential plexus of the molecular layer, at the level of the internal granular and ganglion layers there are two tangential layers of myelin nerve fibers and axonal collaterals of cortical cells. By entering into synaptic connections with cortical neurons, horizontal fibers ensure widespread distribution of nerve impulses in it [5].
Module
I, II, III, IV, V, VI - layers of the cortex Afferent fibers 1. cortico-cortical fiber 2. thalamo-cortical fiber 2a. zone of distribution of specific thalamo-cortical fibers 3. pyramidal neurons 3a. inhibited pyramidal neurons 4. inhibitory neurons and their synapses
4a. cells with an axonal brush 4b. small basket cells 4c. large basket cages 4g. axoaxonal neurons 4d. cells with a double bouquet of dendrites (inhibitory inhibitory neurons) 5. spiny stellate cells, excitatory pyramidal neurons directly and by stimulating cells with a double bouquet of dendrites
Studying the cerebral cortex, J. Szentagothai and representatives of his school established that its structural and functional unit is the module
- vertical column with a diameter of about 300 microns. The module is organized around a cortico-cortical fiber, which is an axon of the pyramidal cell of layer III (layer of pyramidal cells) of the same hemisphere (associative fiber), or from the pyramidal cells of the opposite (commissural). The module includes two thalamo-cortical fibers - specific afferent fibers ending in the fourth layer of the cortex on spiny stellate neurons and extending from the base (basal) dendrites of pyramidal neurons. Each module, according to Szentagothai, is divided into two micromodules with a diameter of less than 100 microns. In total, the human neocortex has approximately 3 million modules. The axons of the pyramidal neurons of the module project to three modules of the same side and through the corpus callosum through commissural fibers to two modules of the opposite hemisphere. In contrast to specific afferent fibers ending in layer IV of the cortex, cortico-cortical fibers form endings in all layers of the cortex, and, reaching layer I, give rise to horizontal branches extending far beyond the module [5].
In addition to specific (thalamo-cortical) afferent fibers, spiny stellate neurons have an excitatory effect on the output pyramidal neurons. There are two types of spiny cells:
- spiny stellate neurons of the focal type
, forming multiple synapses on the dendrites of the pyramidal neuron extending from the apex (apical) - spiny stellate neurons of diffuse type
, the axons of which branch widely in layer IV and excite the basal dendrites of pyramidal neurons. Axon collaterals of pyramidal neurons cause diffuse excitation of neighboring pyramids [5].
The module’s braking system is represented by the following types of neurons:
- cells with an axonal brush
form multiple inhibitory synapses in layer I on the horizontal branches of cortico-cortical fibers - basket neurons
are inhibitory neurons that form inhibitory synapses on the bodies of almost all pyramidal cells. They are divided into small basket neurons, which have an inhibitory effect on pyramidal neurons of layers II, III and V of the module, and large basket cells, located on the periphery of the module and tending to suppress pyramidal neurons of neighboring modules - axoaxonal neurons
, inhibitory pyramidal neurons of layers II and III. Each such cell forms synapses on the initial sections of the axons of hundreds of neurons in layers II and III. They thus inhibit cortico-cortical fibers, but not the projection fibers of layer V neurons [5].
Inhibitory neuron inhibition system:
- cells with a double bouquet of dendrites
are located in layers II and III and, inhibiting inhibitory neurons, produce a secondary excitatory effect on pyramidal neurons. The branches of their axons are directed upward and downward and extend in a narrow column (50 μm). Thus, a cell with a double bouquet of dendrites disinhibits pyramidal neurons in a micromodule (in a column with a diameter of 50-100 μm) [5].
The powerful excitatory effect of focal spiny stellate cells is explained by the fact that they simultaneously excite pyramidal neurons and a cell with a double bouquet of dendrites. Thus, the first three inhibitory neurons inhibit pyramidal cells, and cells with a double bouquet of dendrites excite them, inhibiting inhibitory neurons [5].
However, there are also critical and alternative concepts
, casting doubt on the modular organization of the cerebral cortex and cerebellum. Of course, these views were influenced by the prediction in 1985 and the subsequent discovery in 1992 of diffuse volumetric neurotransmission[6].
Summary
The interneuronal relationships of neurons in the cerebral cortex can be represented as follows: incoming (afferent) information comes from the thalamus along thalamo-cortical fibers, which end on the cells of the IV (internal granular) layer. Its stellate neurons have an excitatory effect on pyramidal cells of layers III (pyramidal neurons) and V (ganglionic) layers, as well as on cells with a double bouquet of dendrites that block inhibitory neurons. Cells of layer III form fibers (associative and commissural) that connect different parts of the cortex. Cells of layers V and VI (multimorphic cells) form projection fibers that extend into the white matter and carry information to other parts of the central nervous system. In all layers of the cortex there are inhibitory neurons that play the role of a filter by blocking pyramidal neurons [5].
The cortex of various sections is characterized by the predominant development of certain layers. Thus, in the motor centers of the cortex, for example, in the anterior central gyrus, layers III, V and VI are highly developed and layers II and IV are poorly expressed. This is the so-called agranular type of bark
.
The descending pathways of the central nervous system originate from these areas. In the sensitive cortical centers, where the afferent conductors coming from the organs of smell, hearing and vision end, the layers containing large and medium pyramidal cells are poorly developed, while the granular layers (II and IV) reach their maximum development. This is a granular type of cortex
[5].
Research methods
Ablations
One of the oldest methods of studying the brain is the ablation technique, which consists of removing one of the parts of the brain, and scientists observe the changes that such an operation leads to.
Not every area of the brain can be removed without killing the organism. Thus, many parts of the brain stem are responsible for vital functions such as breathing, and damage to them can cause immediate death. However, damage to many parts, although it affects the viability of the body, is non-fatal.
Research using ablations has a long history and is currently ongoing. Whereas scientists in the past surgically removed areas of the brain, modern researchers use toxic substances that selectively attack brain tissue (for example, cells in a certain area, but not the nerve fibers passing through it).
After a section of the brain is removed, some functions are lost, while others are retained. For example, a cat whose brain is dissected above the thalamus retains many postural reactions and spinal reflexes. An animal whose brain is dissected at the level of the brainstem (decerebrate) maintains extensor muscle tone but loses postural reflexes.
Observations are also being made of people with lesions of brain structures. Thus, cases of gunshot wounds to the head during the Second World War provided rich information for researchers. Research is also being conducted on patients with stroke and brain damage due to trauma.
Electrophysiology
Electrophysiologists record the electrical activity of the brain - using thin electrodes that allow recording the discharges of individual neurons, or using electroencephalography (a technique for removing brain potentials from the surface of the head).
The thin electrode can be made of metal (coated with an insulating material exposing only the sharp tip) or glass. The glass electrode is a thin tube filled inside with a saline solution. The electrode can be so thin that it penetrates into the cell and allows intracellular potentials to be recorded. Another way to record neuronal activity is extracellular.
In some cases, thin electrodes (from one to several hundred) are implanted into the brain, and researchers record activity over an extended period of time. In other cases, the electrode is inserted into the brain only for the duration of the experiment, and is removed at the end of the recording.
Using a thin electrode, you can record both the activity of individual neurons and local field potentials resulting from the activity of many hundreds of neurons. Using EEG electrodes, as well as surface electrodes placed directly on the brain, it is possible to record only the global activity of a large number of neurons.
When analyzing brain potentials, their spectral analysis is often performed, and different components of the spectrum have different names: delta (0.5 - 4 Hz), theta 1 (4 - 6 Hz), theta 2 (6 - 8 Hz), alpha (8 - 13 Hz ), beta 1 (13 - 20 Hz), beta 2 (20 - 40 Hz), gamma waves (includes the frequency of beta 2 rhythms and higher).
One method for studying brain function is electrical stimulation of specific areas. Using this method, for example, the “motor homunculus” was studied - it was shown that by stimulating certain points in the motor cortex, it is possible to cause arm movement, stimulating other points - leg movements, etc.
If you stimulate the sensory areas of the brain, you can cause sensations. This has been shown both in humans (in the famous Penfield experiments) and in animals.
https://www.youtube.com/watch?v=https:accounts.google.comServiceLogin
Currently, the non-invasive method of focal magnetic stimulation is widely used for brain stimulation. The problem with this method is that it activates quite large areas of the brain, and in some cases it requires stimulation of local areas.
Electrical stimulation is also used in medicine - from electric shock, shown in many films about the horrors of psychiatric clinics, to stimulation of structures deep in the brain, which has become a popular method of treating Parkinson's disease.
Other techniques
X-ray CT and MRI are used to study the anatomical structures of the brain. Also used in anatomical and functional studies of the brain are PET, single-photon emission computed tomography (SPECT), and functional MRI. It is possible to visualize brain structures using ultrasound diagnostics (ultrasound) in the presence of an ultrasound “window” - a defect in the cranial bones, for example, a large fontanel in young children.
Brodmann's cytoarchitectonic fields
Brodmann's cytoarchitectonic areas are sections of the cerebral cortex that differ in their cytoarchitectonics (structure at the cellular level). There are 52 Brodmann cytoarchitectonic areas.
In 1909, the German neurologist Korbinian Brodmann published [7] maps of the cytoarchitectonic fields of the cerebral cortex. Brodmann was the first to create maps of the cortex. Subsequently, O. Vogt and C. Vogt (1919-1920), taking into account the fiber structure, described 150 myeloarchitectonic areas in the cerebral cortex. At the Brain Institute of the USSR Academy of Medical Sciences, I. N. Filippov and S. A. Sarkisov created maps of the cerebral cortex, including 47 cytoarchitectonic fields [8].
Despite criticism [9], Brodmann fields are the best known and most often cited when describing the neuronal organization of the cerebral cortex and its functions.
First and second signaling systems
The role of the cerebral cortex in improving the first signaling system and developing the second is invaluable. These concepts were developed by I.P. Pavlov. The signaling system as a whole is understood as the entire set of processes of the nervous system that carry out perception, processing of information and the response of the body. It connects the body with the outside world.
The first signaling system determines the perception of sensory-specific images through the senses. It is the basis for the formation of conditioned reflexes. This system exists in both animals and humans.
In the higher nervous activity of man, a superstructure has developed in the form of a second signaling system. It is peculiar only to humans and is manifested by verbal communication, speech, and concepts. With the advent of this signaling system, abstract thinking and generalization of countless signals from the first signaling system became possible. According to I.P. Pavlov, words turned into “signals of signals.”
The emergence of the second signaling system became possible thanks to complex labor relationships between people, since this system is a means of communication and collective work. Verbal communication does not develop outside of society. The second signaling system gave rise to abstract (abstract) thinking, writing, reading, counting.
Words are perceived by animals, but completely differently from people. They perceive them as sounds, and not their semantic meaning, like humans. Therefore, animals do not have a second signaling system. Both human signaling systems are interconnected. They organize human behavior in the broad sense of the word. Moreover, the second changed the first signaling system, since the reactions of the first began to largely depend on the social environment. A person has become able to control his unconditioned reflexes, instincts, i.e. first signaling system.
https://www.youtube.com/watch?v=ytcreatorsru
Functions of the cerebral cortex
Familiarity with the most important physiological functions of the cerebral cortex indicates its extraordinary importance in life. The cortex, together with the subcortical formations closest to it, is a department of the central nervous system of animals and humans.
The functions of the cerebral cortex are the implementation of complex reflex reactions that form the basis of higher nervous activity (behavior) of a person. It is no coincidence that it received the greatest development from him. The exclusive properties of the cortex are consciousness (thinking, memory), a second signaling system (speech), and a high organization of work and life in general.
Localization of functions in the cortex
The cerebral cortex functionally consists of three zones: the sensory zone, the motor zone and the association zone. In humans, the association zone occupies about 75% of the cerebral cortex. The function of the association zone is to connect the activity of sensory and motor zones. The association area is thought to receive and process information from the sensory area and initiate goal-directed, meaningful behavior.
Taking action despite fear
, affective, emotionally charged behavior is accompanied by activation of the so-called popliteal region of the anterior cingulate cortex of the brain (subgenual anterior cingulate cortex - sgACC). The greater the fear, the more this area of the brain is activated. At the same time, the activity of the temporal lobes of the brain is suppressed[10].
In July 2020, the journal Nature published information about a map of the cerebral cortex, compiled as a result of research conducted by David Van Essen and his colleagues from the John Washington University School of Medicine. The use of machine learning algorithms made it possible to identify 180 structural areas of the cerebral cortex involved in various functions, occupying 96.6% of the cortex area, including 97 previously unknown ones. As initial data, we used images of multimodal magnetic resonance imaging of the brain of 210 healthy experimental subjects of both sexes who performed simple tasks obtained during the implementation of the project to establish a complete “map” of the structural relationships of the brain “Human Connectome Project” (HCP).[ 11][12]
It was found that the centers of conflict are in the anterior cingulate cortex, and of rejection in the insular lobe. [13]
Neurons for monitoring animals, including other humans, for the purpose of understanding and interacting and synchronizing with them are found in the association parietal (inferior parietal), temporal (superior temporal) and frontal cortices (mirror neurons).
Embryonic development
Embryonic development of the brain is one of the keys to understanding its structure and functions.
The brain develops from the rostral part of the neural tube. Most of the brain (95%) is a derivative of the pterygoid plate.
https://www.youtube.com/watch?v=ytdevru
Embryogenesis of the brain goes through several stages.
- Stage of three brain vesicles - in humans, at the beginning of the fourth week of intrauterine development, the rostral end of the neural tube forms three vesicles: Prosencephalon (forebrain), Mesencephalon (midbrain), Rhombencephalon (diamond-shaped brain, or primary hindbrain).
- Stage of five brain vesicles - in humans, at the beginning of the ninth week of intrauterine development, the Prosencephalon is finally divided into Telencephalon (telecephalon) and Diencephalon (diencephalon), Mesencephalon is preserved, and Rhombencephalon is divided into Metencephalon (hindbrain) and Myelencephalon (medulla oblongata).
During the formation of the second stage (from the third to the seventh weeks of development), the human brain acquires three bends: midbrain, cervical and pontine. First, the midcerebral and pontine flexures are formed simultaneously and in one direction, then the cervical flexure is formed in the opposite direction. As a result, the linear brain “folds” in a zigzag manner.
CGM pathologies and their diagnosis
All disorders of the structure and functioning of the brain and its cortex can be divided into congenital and acquired. Some of these lesions are incompatible with life, for example, anencephaly - complete absence of the brain and acrania - absence of cranial bones.
Other diseases leave a chance for survival, but are accompanied by mental development disorders, for example, encephalocele, in which part of the brain tissue and its membranes protrudes out through an opening in the skull. Microcephaly also falls into this group - an underdeveloped small brain, accompanied by various forms of mental retardation (mental retardation, idiocy) and physical development.
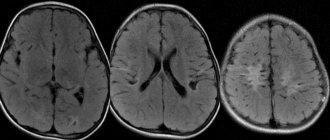
A rarer variant of the pathology is macrocephaly, that is, enlargement of the brain. The pathology is manifested by mental retardation and seizures. With it, the enlargement of the brain can be partial, that is, the hypertrophy is asymmetrical.
Pathologies that affect the cerebral cortex are represented by the following diseases:
- Holoprosencephaly is a condition in which the hemispheres are not separated and there is no complete division into lobes. Children with this disease are stillborn or die within the first day after birth.
- Agyria is underdevelopment of the gyri, in which the functions of the cortex are disrupted. Atrophy is accompanied by multiple disorders and leads to the death of the infant during the first 12 months of life.
- Pachygyria is a condition in which the primary gyri are enlarged to the detriment of the others. The furrows are short and straightened, the structure of the cortex and subcortical structures is disrupted.
- Micropolygyria, in which the brain is covered with small convolutions, and the cortex has not 6 normal layers, but only 4. The condition can be diffuse and local. Immaturity leads to the development of plegia and muscle paresis, epilepsy, which develops in the first year, and mental retardation.
- Focal cortical dysplasia is accompanied by the presence of abnormal areas in the temporal and frontal lobes with huge neurons and abnormal astrocytes. Improper cell structure leads to increased excitability and seizures accompanied by specific movements.
- Heterotopia is an accumulation of nerve cells that during development did not reach their place in the cortex. A single condition can appear after the age of ten; large clusters cause attacks such as epileptic seizures and mental retardation.
Acquired diseases are mainly consequences of serious inflammation, trauma, and also appear after the development or removal of a tumor - benign or malignant. In such conditions, as a rule, the impulse emanating from the cortex to the corresponding organs is interrupted.
The most dangerous is the so-called prefrontal syndrome. This area is actually the projection of all human organs, therefore damage to the frontal lobe leads to disturbances in attention, perception, memory, speech, movements, thinking, as well as partial or complete deformation and changes in the patient’s personality.
A number of pathologies accompanied by external changes or deviations in behavior are quite easy to diagnose, others require more careful study, and removed tumors are subjected to histological examination to exclude a malignant nature.
Alarming indications for the procedure are the presence of congenital pathologies or diseases in the family, fetal hypoxia during pregnancy, asphyxia during childbirth, or birth trauma.
Methods for diagnosing congenital abnormalities
Modern medicine helps prevent the birth of children with severe malformations of the cerebral cortex. To do this, screening is performed in the first trimester of pregnancy, which makes it possible to identify pathologies in the structure and development of the brain at the earliest stages.
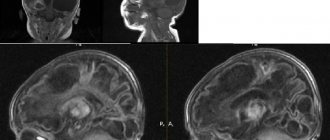
A newborn baby with suspected pathology undergoes neurosonography through the fontanelle, and older children and adults are examined by magnetic resonance imaging. This method allows not only to detect a defect, but also to visualize its size, shape and location.
If there are hereditary problems in the family related to the structure and functioning of the cortex and the entire brain, consultation with a geneticist and specific examinations and tests are required.
The famous “gray cells” are the greatest achievement of evolution and the greatest benefit for humans. Damage can be caused not only by hereditary diseases and injuries, but also by acquired pathologies provoked by the person himself. Doctors urge you to take care of your health, give up bad habits, allow your body and brain to rest and not let your mind get lazy. Loads are useful not only for muscles and joints - they do not allow nerve cells to age and fail. Those who study, work and exercise their brain suffer less from wear and tear and later experience aging and loss of mental abilities.