Нейроны коры больших полушарий головного мозга
Кора больших полушарий головного мозга
или
кора головного мозга
(лат. cortex cerebri) — структура головного мозга, слой серого вещества толщиной 1,3—4,5 мм[1], расположенный по периферии полушарий большого мозга, и покрывающий их. Наибольшая толщина отмечается в верхних участках предцентральной, постцентральной извилин и парацентральной дольки[2].
Кора головного мозга играет очень важную роль в осуществлении высшей нервной (психической) деятельности[2].
У человека кора составляет в среднем 44 % от объёма всего полушария в целом[2]. Площадь поверхности коры одного полушария у взрослого человека в среднем равна 2200 см²[2]. На поверхностные части приходится 1/3, на залегающие в глубине между извилинами — 2/3 всей площади коры[1].
Величина и форма борозд подвержены значительным индивидуальным колебаниям — не только мозг различных людей, но даже полушария одной и той же особи по рисунку борозд не вполне похожи[1].
Всю кору полушарий принято разделять на 4 типа: древняя (палеокортекс), старая (архикортекс), новая (неокортекс) и промежуточная кора (состоящая из промежуточной древней и промежуточной старой коры). Поверхность неокортекса у человека занимает 95,6 %, архикортекса — 2,2 %, палеокортекса — 0,6 %, промежуточной — 1,6 %[2].
Содержание
- 1 Анатомия 1.1 Лобная доля
- 1.2 Теменная доля
- 1.3 Височная доля
- 1.4 Островковая доля (островок)
- 1.5 Борозды и извилины медиальной поверхности
- 1.6 Борозды и извилины нижней поверхности
- 2.1 Строение
Средний мозг
Средний мозг располагается в стволовой части. Он является проводником сигналов от передней части в различные отделы. Его основная функция – регулировка мышечного тонуса. Также он отвечает за передачу тактильных ощущений, координацию и рефлексы. Функции отделов головного мозга человека зависят от их расположения. По этой причине средний мозг отвечает за вестибулярный аппарат. Благодаря среднему мозгу человек может одновременно выполнять несколько функций.
При отсутствии интеллектуальной деятельности нарушается работа мозга. Этому подвержены люди старше 70 лет. При нарушении работы средней части происходят сбои в координации, смещается зрительное и слуховое восприятие.
Анатомия
Доли полушарий большого мозга
Кора большого мозга покрывает поверхность полушарий и образует большое количество различных по глубине и протяжённости борозд (лат. sulci cerebri). Между бороздами расположены различной величины извилины большого мозга (лат. gyri cerebri) [3].
В каждом полушарии различают следующие поверхности:
- выпуклую верхнелатеральную поверхность
(лат. facies superolateralis), примыкающую к внутренней поверхности костей свода черепа - нижнюю поверхность
(лат. facies inferior), передние и средние отделы которой располагаются на внутренней поверхности основания черепа, в области передней и средней черепных ямок, а задние — на намёте мозжечка - медиальную поверхность
(лат. facies medialis), направленную к продольной щели мозга [3].
Эти три поверхности каждого полушария, переходя одна в другую, образуют три края. Верхний край (лат. margo superior) разделяет верхнелатеральную и медиальную поверхности. Нижнелатеральный край (лат. margo inferolateralis) отделяет верхнелатеральную поверхность от нижней. Нижнемедиальный край (лат. margo inferomedialis) располагается между нижней и медиальной поверхностями [3].
В каждом полушарии различают наиболее выступающие места: спереди — лобный полюс (лат. polus frontalis), сзади — затылочный (лат. polus occipitalis), и сбоку — височный (лат. polus temporalis) [3].
Полушарие разделено на пять долей. Четыре из них примыкают к соответствующим костям свода черепа:
- лобная доля (лат. lobus frontalis)
- теменная доля (лат. lobus parietalis)
- затылочная доля (лат. lobus occipitalis)
- височная доля (лат. lobus temporalis)
- островковая доля (лат. lobus insularis) (островок) (лат. insula) — заложена в глубине латеральной ямки большого мозга (лат. fossa lateralis cerebri), отделяющей лобную долю от височной [3].
Лобная доля
Лобная доля
Лобную долю от теменной отделяет глубокая центральная (роландова) борозда (лат. sulcus centralis). Она начинается на медиальной поверхности полушария, переходит на его верхнелатеральную поверхность, идёт по ней немного косо, сзади наперёд, и обычно не доходит до латеральной (боковой или сильвиевой) борозды мозга [3].
Приблизительно параллельно центральной борозде располагается предцентральная борозда (лат. sulcus precentralis), которая не доходит до верхнего края полушария. Предцентральная борозда окаймляет спереди прецентральную извилину (лат. gyrus precentralis) [3].
Верхняя и нижняя лобные борозды (лат. sulci frontales superior et inferior) направляются от предцентральной борозды вперёд. Они делят лобную долю на:
- верхнюю лобную извилину (лат. gyrus frontalis superior), которая расположена выше верхней лобной борозды и переходит на медиальную поверхность полушария
- среднюю лобную извилину (лат. gyrus frontalis medius), которую ограничивают верхняя и нижняя лобные борозды. Орбитальный (передний) сегмент этой извилины переходит на нижнюю поверхность лобной доли
- нижнюю лобную извилину (лат. gyrus frontalis inferior), которая лежит между нижней лобной бороздой и латеральной бороздой мозга и ветвями латеральной борозды делится на ряд частей [3]
Латеральная борозда (лат. sulcus lateralis) — одна из наиболее глубоких борозд головного мозга. Она отделяет височную долю от лобной и теменной. Залегает латеральная борозда на верхнелатеральной поверхности каждого полушария и идёт сверху вниз и кпереди. В глубине этой борозды располагается углубление — латеральная ямка большого мозга (лат. fossa lateralis cerebri), дном которой является наружная поверхность островка[3].
От латеральной борозды к верху отходят мелкие борозды, называемые ветвями. Наиболее постоянными из них являются восходящая (лат. ramus ascendens) и передняя (лат. ramus anterior) ветви. Верхнезадний отдел борозды называется задней ветвью (лат. ramus posterior)[3].
Нижняя лобная извилина, в пределах которой проходят восходящая и передняя ветви, разделяется ими на три части:
- заднюю — покрышечную часть (лат. pars opercularis), ограниченную спереди восходящей ветвью
- среднюю — треугольную часть (лат. pars triangularis), лежащую между восходящей и передней ветвями
- переднюю — глазничную часть (лат. pars orbitalis), расположенные между передней ветвью и нижнелатеральным краем лобной доли[3]
Теменная доля
Теменная доля
Залегает сзади от центральной борозды, которая отделяет её от лобной. От височной отграничена латеральной бороздой мозга, от затылочной — частью теменно-затылочной борозды (лат. sulcus parietooccipitalis)[3].
Параллельно прецентральной извилине проходит постцентральная (лат. gyrus postcentralis). От неё кзади, почти параллельно продольной щели большого мозга, идёт внутритеменная борозда (лат. sulcus intraparietalis), делящая задневерхние отделы теменные отделы теменной доли на две извилины: верхнюю (лат. lobulus parietalis superior) и нижнюю (лат. lobulus parietalis inferior) теменные дольки. В нижней теменной дольке различают две сравнительно небольшие извилины: надкраевую (лат. gyrus supramarginalis), лежащую впереди и замыкающую задние отделы латеральной борозды, и расположенную кзади от предыдущей угловую (лат. gyrus angularis), которая замыкает верхнюю височную борозду[3].
Между восходящей и задней ветвями латеральной борозды мозга расположен участок коры, обозначаемый как лобно-теменная покрышка (лат. operculum frontoparietalis). В неё входят задняя часть нижней лобной извилины, нижние отделы предцентральной и постцентральной извилин, а также нижний отдел передней части теменной доли[3].
Височная доля
Височная доля
Имеет наиболее выраженные границы. В ней различают выпуклую латеральную поверхность и вогнутую нижнюю. Тупой полюс височной доли обращён вперёд и несколько вниз. Латеральная борозда большого мозга резко отграничивает височную долю от лобной[3].
Две борозды, расположенные на верхнелатеральной поверхности: верхняя (лат. sulcus temporalis superior) и нижняя (лат. sulcus temporalis inferior) височные борозды, следуя почти параллельно латеральной борозде мозга, разделяют долю на три височные извилины: верхнюю, среднюю и нижнюю (лат. gyri temporales superior, medius et inferior)[3].
Те участки височной доли, которые направлены в сторону латеральной борозды мозга изрезаны короткими поперечными височными бороздами (лат. sulci temporales transversi). Между этими бороздами залегают 2-3 короткие поперечные височные извилины, связанные с извилинами височной доли (лат. gyri temporales transversi) и островком[3].
Островковая доля (островок)
Залегает на дне латеральной ямки большого мозга (лат. fossa lateralis cerebri).
Она представляет собой трёхстороннюю пирамиду, обращённую своей вершиной — полюсом островка — кпереди и кнаружи, в сторону латеральной борозды. С периферии островок окружён лобной, теменной и височной долями, участвующими в образовании стенок латеральной борозды мозга[3].
Основание островка с трёх сторон окружено круговой бороздой островка (лат. sulcus circularis insulae).
Его поверхность прорезана глубокой центральной бороздой островка (лат. sulcus centralis insulae). Эта борозда разделяет островок на переднюю и заднюю части[3].
На поверхности различают большое количество мелких извилин островка (лат. gyri insulae). Большая передняя часть состоит из нескольких коротких извилин островка (лат. gyri breves insulae), задняя — одной длинной извилины (лат. gyrus longus insulae)[3].
Борозды и извилины медиальной поверхности
На медиальную поверхность полушария выходят лобная, теменная и затылочная доли.
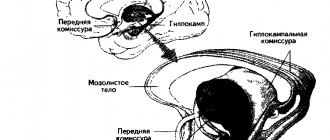
Поясная извилина (лат. gyrus cinguli) начинается подмозолистым полем (лат. area subcallosa), огибает мозолистое тело и при посредстве узкой полоски — перешейка поясной извилины (лат. isthmus gyri cinguli) переходит в парагиппокампальную извилину на нижней поверхности полушария[3].
Борозда мозолистого тела (лат. g sulcus corporis callosi) отделяет поясную извилину от мозолистого тела и на нижней поверхности полушария продолжается в борозду гиппокампа[3].
Поясная извилина ограничена сверху поясной бороздой (лат. sulcus cinguli). В последней различают выпуклую по направлению к лобному полюсу переднюю часть и заднюю часть, которая, следуя вдоль поясной извилины и не доходя до её заднего отдела, поднимается к верхнему краю полушария большого мозга. Задний конец борозды лежит позади верхнего конца центральной борозды. Между предцентральной бороздой, окончание которой иногда хорошо видно у верхнего края медиальной поверхности полушария, и концом поясной борозды, располагается парацентральная долька (лат. lobulus paracentralis)[3].
Выше поясной извилины, Спереди от подмозолистого поля, начинается медиальная лобная извилина (лат. gyrus frontalis medialis). Она тянется до парацентральной дольки и является нижней частью верхней лобной извилины.
Сзади от поясной борозды лежит небольшая четырёхугольная долька — предклинье (лат. precuneus). Её задней границей является глубокая теменно-затылочная борозда (лат. sulcus parietooccipitalis), нижней — подтеменная борозда (лат. sulcus subparietalis), отделяющая предклинье от заднего отдела поясной извилины[3].
Сзади и ниже предклинья залегает треугольная долька — клин (лат. cuneus). Выпуклая наружная поверхность клина участвует в образовании затылочного полюса. Направленная вниз и вперёд вершина клина почти доходит до заднего отдела поясной извилины. Задненижней границей клина является очень глубокая шпорная борозда (лат. sulcus calcarinus), передней — теменно-затылочная борозда[3].
Борозды и извилины нижней поверхности
На нижней поверхности лобной доли располагается обонятельная борозда (лат. sulcus olfactorius). Кнутри от неё, между нею и нижнемедиальным краем полушария, лежит прямая извилина (лат. gyrus rectus). Её задний отдел доходит до переднего продырявленного вещества (лат. substantia perforata anterior). Кнаружи от борозды располагается остальная часть нижней поверхности лобной доли, изрезанная короткими глазничными бороздами (лат. sulci orbitales), на ряд небольших глазничных извилин (лат. gyri orbitales)[3].
Нижняя поверхность височной доли глубокой бороздой гиппокампа (лат. sulcus hippocampi) отделена от ножек мозга. В глубине борозды залегает узкая зубчатая извилина (лат. gyrus dentatus). Передний её конец переходит в крючок, а задний — в ленточную извилину (лат. gyrus fasciolaris) залегающую под валиком мозолистого тела. Латерально от борозды находится парагиппокампальная извилина (лат. gyrus parahippocampalis). Впереди эта извилина имеет утолщение в виде крючка (лат. uncus), а кзади продолжается в язычную извилину (лат. gyrus lingualis). Парагиппокампальную и язычную извилины с латеральной стороны ограничивает коллатеральная борозда (лат. sulcus collateralis), переходящая спереди в носовую борозду (лат. sulcus rhinalis). Остальную часть нижней поверхности височной доли занимают медиальная и латеральная затылочно-височные извилины (лат. gyri occipitotemporales medialis et lateralis), разделённые затылочно-височной бороздой (лат. sulcus occipitotemporalis). Латеральная затылочно-височная извилина нижнелатеральным краем полушария отделяется от нижней височной извилины[3].
Функции полушарий
Правое и левое полушария у человека выполняют разные функции. В левом полушарии находятся центры устной и письменной речи. Здесь же осуществляются процессы анализа и синтеза информации, делаются обобщения и принимаются решения. Обеспечиваемое левым полушарием словесно-логическое мышление позволяет познать сущность объекта, выйти за пределы индивидуального мира. На его основе формируется человеческое знание. Оно передается из поколения в поколение посредством записи словесных или знаковых сигналов.
Правое полушарие осуществляет образное мышление. Оперируя образами объектов внешнего мира, оно может создавать из них небывалые, фантастические комбинации. А это — основа творчества, принятия необычных решений. Исключительно велико значение правого полушария для музыкального и художественного творчества. Известно, что наиболее выдающиеся художники, поэты, музыканты — это люди с преобладанием правополушарного мышления.
Несмотря на функциональную асимметрию, мозг работает как единое целое. Обобщая информацию, он обеспечивает адекватное поведение, мышление, сознание, память, трудовую и творческую деятельность человека.
Гистология
Строение
Цитоархитектоника (расположение клеток)
- молекулярный слой
- наружный зернистый слой
- слой пирамидальных нейронов
- внутренний зернистый слой
- ганглионарный слой (внутренний пирамидный слой;клетки Беца)
- слой полиморфных клеток
Миелоархитектоника (расположение волокон)
- полоска молекулярного слоя
- полоска наружного зернистого слоя
- полоска внутреннего зернистого слоя
- полоска ганглионарного слоя [4].
Кора полушарий головного мозга представлена слоем серого вещества толщиной в среднем около 3 мм (1,3 — 4,5 мм). Наиболее сильно развита она в передней центральной извилине. Обилие борозд и извилин значительно увеличивает площадь серого вещества головного мозга. В коре содержится около 10-14 млрд нервных клеток. Различные её участки, отличающиеся друг от друга некоторыми особенностями расположения и строения клеток (цитоархитектоника), расположения волокон (миелоархитектоника) и функциональным значением, называются полями. Они представляют собой места высшего анализа и синтеза нервных импульсов. Резко очерченные границы между ними отсутствуют. Для коры характерно расположение клеток и волокон слоями [5].
Типичным для новой коры (лат. neocortex) является наличие шести слоёв, различающихся между собой главным образом по форме входящих в них нервных клеток. При этом на медиальной и нижней поверхностях полушарий сохранились участки старой (лат. archipallium) и древней (лат. paleopallium) коры, имеющей 2-слойное и 3-слойное строение[1]. Также выделяется промежуточная кора (лат. mesopallium) располагающаяся между старой и новой, а также древней и новой корой[3]. Древняя кора представлена гиппокампом, а старая — участком коры возле обонятельной луковицы на нижней поверхности лобной доли[1].
Цитоархитектоника
Мультиполярные нейроны коры головного мозга весьма разнообразны по форме. Среди них можно выделить:
- пирамидные
- звёздчатые
- веретенообразные
- паукообразные
- горизонтальные
Пирамидные нейроны составляют основную и наиболее специфическую для коры головного мозга форму (80—90 % всех нейронов). Размеры их варьируют от 10 до 140 мкм. Они имеют вытянутое треугольное тело, вершина которого обращена к поверхности коры. От вершины и боковых поверхностей тела отходят дендриты, заканчивающиеся в различных слоях серого вещества. От основания пирамидных клеток берут начало аксоны, в одних клетках короткие, образующие ветвления в пределах данного участка коры, в других — длинные, поступающие в белое вещество [5].
Пирамидные клетки различных слоёв коры отличаются размерами и имеют разное функциональное значение. Мелкие клетки представляют собой вставочные нейроны, аксоны которых связывают отдельные участки коры одного полушария (ассоциативные нейроны) или двух полушарий (комиссуральные нейроны). Эти клетки встречаются в разных количествах во всех слоях коры. Особенно богата ими кора головного мозга человека. Аксоны крупных пирамидных нейронов принимают участие в образовании пирамидных путей, проецирующих импульсы в соответствующие центры мозгового ствола и спинного мозга [5].
Нейроны коры расположены нерезко отграниченными слоями. Каждый слой характеризуется преобладанием какого-либо одного вида клеток. В двигательной зоне коры различают 6 основных слоёв:
- Молекулярный (лат. lamina molecularis)
- Наружный зернистый (лат. lamina granularis externa)
- Пирамидальных нейронов (лат. lamina pyramidalis)
- Внутренний зернистый (лат. lamina granularis interna)
- Ганглионарный (слой клеток Беца) (лат. lamina ganglionaris)
- Слой мультиформных (полиморфных) клеток (лат. lamina multiformis) [5]
Кора полушарий головного мозга также содержит мощный нейроглиальный аппарат, выполняющий трофическую, защитную, опорную и разграничительную функции [5].
На медиальной и нижней поверхности полушарий сохранились участки старой, древней коры, которые имеют двухслойное и трехслойное строение.
Молекулярный слой
Молекулярный слой коры содержит небольшое количество мелких ассоциативных клеток веретеновидной формы. Их аксоны проходят параллельно поверхности мозга в составе тангенциального сплетения нервных волокон молекулярного слоя. Основная масса волокон этого сплетения представлена ветвлениями дендритов нейронов нижележащих слоёв [5].
Наружный зернистый слой
Наружный зернистый слой образован мелкими нейронами диаметром около 10 мкм, имеющими округлую, угловатую и пирамидальную форму, и звёздчатыми нейронами. Дендриты этих клеток поднимаются в молекулярный слой. Аксоны или уходят в белое вещество, или, образуя дуги, также поступают в тангенциальное сплетение волокон молекулярного слоя [5].
Слой пирамидальных нейронов
Является самым широким по сравнению с другими слоями коры головного мозга. Он особенно хорошо развит в прецентральной извилине. Величина пирамидных клеток последовательно увеличивается в пределах 10-40 мкм от наружной зоны этого слоя к внутренней. От верхушки пирамидной клетки отходит главный дендрит, который располагается в молекулярном слое. Дендриты, берущие начало от боковых поверхностей пирамиды и её основания, имеют незначительную длину и образуют синапсы со смежными клетками этого слоя. Аксон пирамидной клетки всегда отходит от её основания. В мелких клетках он остаётся в пределах коры; аксон же, принадлежащий крупной пирамиде, обычно формирует миелиновое ассоцативное или комиссуральное волокно, идущее в белое вещество [5].
Внутренний зернистый слой
В некоторых полях коры развит очень сильно (например, в зрительной зоне коры). Однако в других участках он может отсутствовать (в прецентральной извилине). Этот слой образован мелкими звёздчатыми нейронами. В его состав входит большое количество горизонтальных волокон [5].
Ганглионарный слой (Внутренний пирамидный слой;Клетки Беца)
Образован крупными пирамидными клетками, причём область прецентральной извилины содержит гигантские клетки, описанные впервые российским анатомом В. А. Бецем в 1874 году (клетки Беца). Они достигают в высоту 120 и в ширину 80 мкм. В отличие от других пирамидных клеток коры гигантские клетки Беца характеризуются наличием крупных глыбок хроматофильного вещества. Их аксоны образуют главную часть кортико-спинальных и кортико-нуклеарных путей и оканчиваются на мотонейронах мозгового ствола и спинного мозга [5].
Перед выходом из коры от пирамидного пути отходит множество коллатералей. Аксоны от гигантских клеток Беца дают коллатерали, посылающие тормозящие импульсы в саму кору. Также коллатерали волокон пирамидного пути идут в полосатое тело, красное ядро, ретикулярную формацию, ядра моста и нижних олив. Ядра моста и нижних олив передают сигнал в мозжечок. Таким образом, когда пирамидный путь передаёт сигнал, вызывающий целенаправленное движение, в спинной мозг, одновременно сигналы получают базальные ганглии, ствол мозга и мозжечок. Помимо коллатералей пирамидных путей, существуют волокна, которые идут непосредственно от коры к промежуточным ядрам: хвостатому телу, красному ядру, ядрам ретикулярной формации ствола мозга и др.[5]
Слой мультиморфных клеток
Образован нейронами различной, преимущественно веретенообразной формы. Внешняя зона этого слоя содержит более крупные клетки. Нейроны внутренней зоны мельче и лежат на большом расстоянии друг от друга. Аксоны клеток полиморфного слоя уходят в белое вещество в составе эфферентных путей головного мозга. Дендриты достигают молекулярного слоя коры [5].
Миелоархитектоника
Среди нервных волокон коры полушарий головного мозга можно выделить:
- ассоциативные волокна — связывают отдельные участки коры одного полушария
- комиссуральные волокна — соединяют кору двух полушарий
- проекционные волокна — соединяют кору с ядрами низших отделов центральной нервной системы. Афферентные проекционные волокна заканчиваются в слое пирамидальных нейронов[5]
Кроме тангенциального сплетения молекулярного слоя, на уровне внутреннего зернистого и ганглионарного слоёв расположены два тангенциальных слоя миелиновых нервных волокон и коллатералей аксонов клеток коры. Вступая в синаптические связи с нейронами коры, горизонтальные волокна обеспечивают широкое распространение в ней нервного импульса [5].
Модуль
I, II, III, IV, V, VI — слои коры Афферентные волокна 1. кортико-кортикальное волокно 2. таламо-кортикальное волокно 2а. зона распространения специфических таламо-кортикальных волокон 3. пирамидные нейроны 3а. заторможенные пирамидные нейроны 4. тормозные нейроны и их синапсы
4а. клетки с аксональной кисточкой 4б. малые корзинчатые клетки 4в. большие корзинчатые клетки 4г. аксоаксональные нейроны 4д. клетки с двойным букетом дендритов (тормозящие тормозные нейроны) 5. шипиковые звёздчатые клетки, возбуждающие пирамидные нейроны непосредственно и путём стимуляции клеток с двойным букетом дендритов
Исследуя кору больших полушарий головного мозга Я. Сентаготаи и представители его школы установили, что её структурно-функциональной единицей является модуль
— вертикальная колонка диаметром около 300 мкм. Модуль организован вокруг кортико-кортикального волокна, представляющего собой аксон пирамидной клетки III слоя (слоя пирамидальных клеток) того же полушария (ассоциативное волокно), либо от пирамидальных клеток противоположного (комиссуральное). В модуль входят два таламо-кортикальных волокна — специфических афферентных волокна, оканчивающихся в IV слое коры на шипиковых звёздчатых нейронах и отходящих от основания (базальных) дендритах пирамидальных нейронов. Каждый модуль, по мнению Сентаготаи разделяется на два микромодуля диаметром менее 100 мкм. Всего в неокортексе человека примерно 3 млн модулей. Аксоны пирамидальных нейронов модуля проецируются на три модуля той же стороны и через мозолистое тело посредством комиссуральных волокон на два модуля противоположного полушария. В отличие от специфических афферентных волокон, оканчивающихся в IV слое коры, кортико-кортикальные волокна образуют окончания во всех слоях коры, и, достигая I слоя, дают горизонтальные ветви, выходящие далеко за пределы модуля[5].
Помимо специфических (таламо-кортикальных) афферентных волокон, на выходные пирамидальные нейроны возбуждающее влияние оказывают шипиковые звёздчатые нейроны. Различают два типа шипиковых клеток:
- шипиковые звёздчатые нейроны фокального типа
, образующие множественные синапсы на отходящих от верхушки (апикальных) дендритах пирамидального нейрона - шипиковые звёздчатые нейроны диффузного типа
, аксоны которых широко ветвятся в IV слое и возбуждают базальные дендриты пирамидальных нейронов. Коллатерали аксонов пирамидных нейронов вызывают диффузное возбуждение соседних пирамид [5].
Тормозная система модуля представлена следующими типами нейронов:
- клетки с аксональной кисточкой
образуют в I слое множественные тормозные синапсы на горизонтальных ветвях кортико-кортикальных волокон - корзинчатые нейроны
— тормозные нейроны, образующие тормозящие синапсы на телах практически всех пирамидных клеток. Они подразделяются на малые корзинчатые нейроны, оказывающие тормозящее влияние на пирамидные нейроны II, III и V слоёв модуля, и большие корзинчатые клетки, располагающиеся на периферии модуля и имеющие тенденцию подавлять пирамидные нейроны соседних модулей - аксоаксональные нейроны
, тормозящие пирамидные нейроны II и III слоёв. Каждая такая клетка образует синапсы на начальных участках аксонов сотен нейронов II и III слоёв. Они тормозят, таким образом, кортико-кортикальные волокна, но не проекционные волокна нейронов V слоя[5].
Система угнетения тормозных нейронов:
- клетки с двойным букетом дендритов
располагаются во II и III слоях и, угнетая тормозные нейроны, производят вторичное возбуждающее действие на пирамидные нейроны. Ветви их аксонов направлены вверх и вниз и распространяются в узкой колонке (50 мкм). Таким образом, клетка с двойным букетом дендритов растормаживает пирамидные нейроны в микромодуле (в колонке диаметром 50-100 мкм)[5].
Мощный возбуждающий эффект фокальных шипиковых звёздчатых клеток объясняется тем, что они одновременно возбуждают пирамидные нейроны и клетку с двойным букетом дендритов. Таким образом, первые три тормозных нейрона тормозят пирамидные клетки, а клетки с двойным букетом дендритов возбуждают их, угнетая тормозные нейроны[5].
Однако, также существуют критические и альтернативные концепции
, ставящие под сомнение модульную организацию коры больших полушарий и мозжечка. Безусловно, влияние на эти воззрения оказало предсказание в 1985 г. и в дальнейшем открытие в 1992 г. диффузного объёмного нейротрансмиттинга[6].
Резюме
Межнейрональные взаимосвязи нейронов коры больших полушарий головного мозга можно представить следующим образом: входящая (афферентная) информация поступает из таламуса по таламо-кортикальным волокнам, которые заканчиваются на клетках IV (внутреннего зернистого) слоя. Его звёздчатые нейроны оказывают возбуждающее воздействие на пирамидные клетки III (пирамидальных нейронов) и V (ганглионарного) слоёв, а также на клетки с двойным букетом дендритов, которые блокируют тормозные нейроны. Клетки III слоя образуют волокна (ассоциативные и комиссуральные), которые связывают между собой различные отделы коры. Клетки V и VI (мультиморфных клеток) слоёв формируют проекционные волокна, которые уходят в белое вещество и несут информацию другим отделам центральной нервной системы. Во всех слоях коры находятся тормозные нейроны, играющие роль фильтра путём блокирования пирамидных нейронов [5].
Кора различных отделов характеризуется преимущественным развитием тех или иных её слоёв. Так, в двигательных центрах коры, например в передней центральной извилине, сильно развиты III, V и VI и плохо выражены II и IV слои. Это так называемый агранулярный тип коры
. Из этих областей берут начало нисходящие проводящие пути центральной нервной системы. В чувствительных корковых центрах, где заканчиваются афферентные проводники, идущие от органов обоняния, слуха и зрения, слабо развиты слои, содержащие крупные и средние пирамидные клетки , тогда как зернистые слои (II и IV) достигают своего максимального развития. Это
гранулярный тип коры
[5].
Методы исследования
Аблации
Одним из старейших методов исследования мозга является методика аблаций, которая состоит в том, что один из отделов мозга удаляется, и ученые наблюдают за изменениями, к которым приводит такая операция.
Не всякую область мозга можно удалить, не убив организм. Так, многие отделы ствола мозга ответственны за жизненно важные функции, такие, как дыхание, и их поражение может вызвать немедленную смерть. Тем не менее, поражение многих отделов, хотя и отражается на жизнеспособности организма, несмертельно.
Исследования с применением аблаций имеют давнюю историю и продолжаются в настоящее время. Если ученые прошлого удаляли области мозга хирургическим путем, то современные исследователи используют токсические вещества, избирательно поражающие ткани мозга (например, клетки в определенной области, но не проходящие через нее нервные волокна).
После удаления отдела мозга какие-то функции теряются, а какие-то сохраняются. Например, кошка, мозг которой рассечен выше таламуса, сохраняет многие позные реакции и спинномозговыерефлексы. Животное, мозг которого рассечен на уровне ствола мозга (децеребрированное), поддерживает тонус мышц-разгибателей, но утрачивает позные рефлексы.
Проводятся наблюдения и за людьми с поражениями мозговых структур. Так, богатую информацию для исследователей дали случаи огнестрельных ранений головы во время Второй Мировой войны. Также проводятся исследования больных, пораженных инсультом, и с поражениями мозга в результате травмы.
Электрофизиология
Электрофизиологи регистрируют электрическую активность мозга — с помощью тонких электродов, позволяющих записывать разряды отдельных нейронов, или с помощью электроэнцефалографии (методики отведения потенциалов мозга с поверхности головы).
Тонкий электрод может быть сделан из металла (покрытого изоляционным материалом, обнажающим лишь острый кончик) или из стекла. стеклянный электрод представляет собой тонкую трубочку, заполненную внутри солевым раствором. Электрод может быть настолько тонок, что проникает внутрь клетки и позволяет записывать внутриклеточные потенциалы. Другой способ регистрации активности нейронов — внеклеточный.
В некоторых случаях тонкие электроды (от одного до несколько сотен) вживляются в мозг, и исследователи регистрируют активность продолжительное время. В других случаях электрод вводится в мозг только на время эксперимента, а по окончании записи извлекается.
С помощью тонкого электрода можно регистрировать как активность отдельных нейронов, так локальные потенциалы (local field potentials), образующиеся в результате активности многих сотен нейронов. С помощью ЭЭГ электродов, а также поверхностных электродов, накладываемых непосредственно на мозг, можно регистрировать только глобальную активность большого количества нейронов.
При анализе потенциалов мозга часто производят их спектральный анализ, причем разные компоненты спектра имеют разные названия: дельта (0.5 — 4 Гц), тета 1 (4 — 6 Гц), тета 2 (6 — 8 Гц), альфа (8 — 13 Гц), бета 1 (13 — 20 Гц), бета 2 (20 — 40 Гц), гамма волны (включает частоту бета 2 ритма и выше).
Одним из методов изучения функций мозга является электрическая стимуляция отдельных областей. С помощью этого метода был, например, исследован «моторный гомункулус» — было показано, что, стимулируя определенные точки в моторной коре, можно вызвать движение руки, стимулируя другие точки — движения ног и т. д.
Если же стимулировать сенсорные области мозга, то можно вызвать ощущения. Это было показано как на человеке (в знаменитых опытах Пенфилда), так и на животных.
https://www.youtube.com/watch?v=https:accounts.google.comServiceLogin
В настоящее время для стимуляции мозга широко используется неинвазивный метод фокальной магнитной стимуляции. Проблема с этим методом состоит в том, что он активирует довольно большие участки мозга, а в некоторых случаях требуется стимулировать локальные участки.
Применяется электрическая стимуляция и в медицине — от электрошока, показанного во многих кинофильмах об ужасах психиатрических клиник, до стимуляции структур в глубине мозга, ставшей популярным методом лечения болезни Паркинсона.
Другие методики
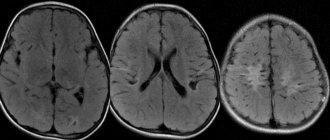
Для исследования анатомических структур головного мозга применяются рентгеновская КТ и МРТ. Также при анатомо-функциональных исследованиях головного мозга применяются ПЭТ, однофотонная эмиссионная компьютерная томография (ОФЭКТ), функциональная МРТ. Возможна визуализация структур головного мозга методом ультразвуковой диагностики (УЗИ) при наличии ультразвукового «окна» — дефекта черепных костей,например, большой родничок у детей раннего возраста.
Цитоархитектонические поля Бродмана
Цитоархитектонические поля Бродмана – отделы коры больших полушарий головного мозга, отличающиеся по своей цитоархитектонике (строению на клеточном уровне). Выделяется 52 цитоархитектонических поля Бродмана.
В 1909 году немецкий невролог Корбиниан Бродманн опубликовал [7] карты цитоархитектонических полей коры больших полушарий головного мозга. Бродман впервые создал карты коры. Впоследствии О. Фогт и Ц. Фогт (1919-1920 гг.) с учётом волоконного строения описали в коре головного мозга 150 миелоархитектонических участков. В Институте мозга АМН СССР И. Н. Филипповым и С. А. Саркисовым были созданы карты коры головного мозга, включающие 47 цитоархитектонических полей[8].
Несмотря на критику [9], поля Бродмана являются самыми известными и наиболее часто цитируемыми при описании нейрональной организации коры головного мозга и её функций.
Первая и вторая сигнальные системы
Неоценима роль коры большого мозга в совершенствовании первой сигнальной системы и развитии второй. Эти понятия разработаны И.П.Павловым. Под сигнальной системой в целом понимают всю совокупность процессов нервной системы, осуществляющих восприятие, переработку информации и ответную реакцию организма. Она связывает организм с внешним миром.
Первая сигнальная система обусловливает восприятие посредством органов чувств чувственно-конкретных образов. Она является основой для образования условных рефлексов. Эта система существует как у животных, так и у человека.
В высшей нервной деятельности человека развилась надстройка в виде второй сигнальной системы. Она свойственна только человеку и проявляется словесным общением, речью, понятиями. С появлением этой сигнальной системы стали возможными отвлеченное мышление, обобщение бесчисленных сигналов первой сигнальной системы. По И.П.Павлову, слова превратились в «сигналы сигналов».
Возникновение второй сигнальной системы стало возможным благодаря сложным трудовым взаимоотношениям между людьми, так как эта система является средством общения, коллективного труда. Словесное общение не развивается вне общества. Вторая сигнальная система породила отвлеченное (абстрактное) мышление, письмо, чтение, счет.
Слова воспринимаются и животными, но совершенно отлично от людей. Они воспринимают их как звуки, а не их смысловое значение, как люди. Следовательно, у животных нет второй сигнальной системы. Обе сигнальные системы человека взаимосвязаны. Они организуют поведение человека в широком смысле слова. Причем вторая изменила первую сигнальную систему, так как реакции первой стали в значительной мере зависеть от социальной среды. Человек стал в состоянии управлять своими безусловными рефлексами, инстинктами, т.е. первой сигнальной системой.
https://www.youtube.com/watch?v=ytcreatorsru
Функции коры мозга
Знакомство с наиболее важными физиологическими функциями коры большого мозга свидетельствует о необычайном ее значении в жизнедеятельности. Кора вместе с ближайшими к ней подкорковыми образованиями является отделом центральной нервной системы животных и человека.
Функции коры головного мозга — осуществление сложных рефлекторных реакций, составляющих основу высшей нервной деятельности (поведения) человека. Не случайно у него она получила наибольшее развитие. Исключительным свойством коры являются сознание (мышление, память), вторая сигнальная система (речь), высокая организация труда и жизни в целом.
Локализация функций в коре
Кора головного мозга функционально состоит из трех зон: сенсорная зона, моторная зона и ассоциативная зона. У человека ассоциативная зона занимает около 75% коры головного мозга. Функция ассоциативной зоны — связывать между собой активность сенсорных и моторных зон. Ассоциативная зона, предполагается, получает и перерабатывает информацию из сенсорной зоны и инициирует целенаправленное осмысленное поведение.
Действия вопреки страху
, аффективное, эмоционально насыщенное поведение сопровождаются активацией так называемой подколенной области передней части поясной извилины головного мозга (subgenual anterior cingulate cortex — sgACC). Чем больше страх, тем сильнее активизируется эта область мозга. При этом одновременно подавляляется активность височных долей головного мозга[10].
В июле 2020 на сайте журнала Nature были опубликована информация о карте коры головного мозга, составленной в результате исследований, проведенных Дэвидом Ван Эссеном (David Van Essen) и его коллегами из Медицинской школы Университета Дж. Вашингтона. Использование алгоритмов машинного обучения позволило идентифицировать 180 структурных участков коры головного мозга, вовлеченных в выполнение различных функций, занимающих 96.6% площади коры, включая 97 прежде неизвестных. В качестве исходных данных использовались изображения мультимодальной магнитно-резонансной томографии головного мозга 210 здоровых подопытных обоих полов, выполнявших простые задания, полученные в ходе реализации проекта по установлению полной «карты» структурных взаимосвязей мозга «Коннектом человека» (Human Connectome Project, HCP).[11][12]
Обнаружено, что центры конфликта находятся в передней поясной коре, неприятия в островковой доле. [13]
Нейроны наблюдения за животными, в том числе другими людьми, с целью понимания и обеспечения взаимодействия и синхронизации с ними, найдены в ассоциативной теменной (нижняя теменная), височной (верхняя височная) и лобной коре (зеркальные нейроны).
Эмбриональное развитие
Эмбриональное развитие мозга является одним из ключей к пониманию его строения и функций.
Головной мозг развивается из ростральной части нервной трубки. Большая часть головного мозга (95 %) является производной крыловидной пластинки.
https://www.youtube.com/watch?v=ytdevru
Эмбриогенез мозга проходит через несколько стадий.
- Стадия трех мозговых пузырей — у человека в начале четвертой недели внутриутробного развития ростральный конец нервной трубки формирует три пузыря: Prosencephalon (передний мозг), Mesencephalon (средний мозг), Rhombencephalon (ромбовидный мозг, или первичный задний мозг).
- Стадия пяти мозговых пузырей — у человека в начале девятой недели внутриутробного развития Prosencephalon окончательно делится на Telencephalon (конечный мозг) и Diencephalon (промежуточный мозг), Mesencephalon сохраняется, а Rhombencephalon делится на Metencephalon (задний мозг) и Myelencephalon (продолговатый мозг).
В процессе формирования второй стадии (с третьей по седьмую недели развития) головной мозг человека приобретает три изгиба: среднемозговой, шейный и мостовый. Сначала одновременно и в одном направлении формируются среднемозговой и мостовый изгибы, потом — и в противоположном направлении — шейный. В итоге линейный мозг зигзагообразно «складывается».
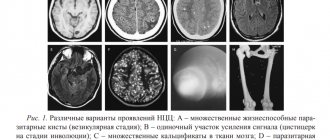
Патологии КГМ и их диагностика
Все нарушения строения и функционирования головного мозга и его коры можно разделить на врожденные и приобретенные. Часть из таких поражений несовместима с жизнью, например, анэнцефалия – полное отсутствие мозга и акрания – отсутствие черепных костей.
Другие заболевания оставляют шанс на выживание, но сопровождаются нарушениями умственного развития, например, энцефалоцеле, при котором часть мозговых тканей и его оболочек выпячивается наружу через отверстие в черепе. В эту же группу попадает и микроцефалия – недоразвитый маленький мозг, сопровождающийся разными формами задержки психического (олигофрения, идиотия) и физического развития.
Более редким вариантом патологии является макроцефалия, то есть увеличение головного мозга. Патология проявляется умственной отсталостью и судорогами. При нем увеличение мозга может быть частичным, то есть гипертрофия асимметричная.
Патологии, при которых поражается кора головного мозга, представлены следующими заболеваниями:
- Голопрозэнцефалия – состояние, при котором полушария не разделены и не существует полноценного деления на доли. Дети при такой болезни рождаются мертвыми или погибают в первые сутки после родов.
- Агирия – недоразвитость извилин, при котором нарушаются функции коры. Атрофия сопровождается множественными расстройствами и приводит к смерти младенца в течение первых 12 месяцев жизни.
- Пахигирия – состояние, при котором первичные извилины увеличены в ущерб остальным. Борозды при этом короткие и выпрямленные, строение коры и подкорковых структур нарушено.
- Микрополигирия, при которой мозг покрыт мелкими извилинами, а кора имеет не 6 нормальных слоев, а всего 4. Состояние бывает диффузным и локальным. Незрелость приводит к развитию плегий и парезов мышц, эпилепсии, которая развивается в первый же год, умственной отсталости.
- Фокальная корковая дисплазия сопровождается наличием в височной и лобной доле патологических участков с огромными нейронами и ненормальными астроцитами. Неправильное строение клеток приводит к возникновению повышенной возбудимости и приступам, сопровождающимся специфическими движениями.
- Гетеротопия – скопление нервных клеток, которые в процессе развития не достигли своего места в коре. Одиночное состояние может проявиться после десятилетнего возраста, большие скопления вызывают приступы типа эпилептических припадков и олигофрению.
Приобретенные заболевания в основном являются следствиями перенесенных серьезных воспалений, травм, а также появляются после развития или удаления опухоли – доброкачественной или злокачественной. При таких состояниях, как правило, прерывается импульс, исходящий от коры в соответствующие органы.
Наиболее опасным считается так называемый префронтальный синдром. Эта область – фактически проекция всех органов человека, поэтому повреждения лобной доли приводит к нарушениям внимания, восприятия, памяти, речи, движений, мышления, а также к частичной или полной деформации и изменению личности больного.
Ряд патологий, сопровождающихся внешними изменениями или отклонениями в поведении, диагностировать достаточно легко, другие требуют более тщательного изучения, а удаленные опухоли подвергаются гистологическому исследованию, чтобы исключить злокачественную природу.
Тревожными показаниями для проведения процедуры является наличие в семье врожденных патологий или заболеваний, гипоксия плода в беременности, асфиксия в родах, родовая травма.
Методы диагностики врожденных отклонений
Современная медицина помогает препятствовать рождению детей с тяжелейшими пороками развития коры головного мозга. Для этого выполняется скрининг в первом триместре беременности, который позволяет выявить патологии строения и развития мозга на самых ранних стадиях.
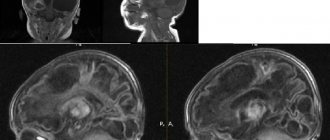
У родившегося крохи с подозрением на патологии проводится нейросонография через «родничок», а детей постарше и взрослых обследуют путем проведения магнитно-резонансной томографии. Этот способ позволяет не только обнаружить дефект, но и визуализировать его размеры, форму и расположение.
Если в семье встречались наследственные проблемы, связанные со строением и функционированием коры и всего мозга, требуется консультация генетика и проведение специфических обследований и анализов.
Знаменитые «серые клеточки» – величайшее достижение эволюции и высшее благо для человека. Вызвать повреждения могут не только наследственные заболевания и травмы, но и приобретенные патологии, спровоцированные самим человеком. Врачи призывают беречь здоровье, отказаться от вредных привычек, позволять своему телу и мозгу отдыхать и не давать разуму лениться. Нагрузки полезны не только мышцам и суставам – они не позволяют нервным клеткам стареть и выходить из строя. Тот, кто учится, работает и загружает свой мозг, меньше страдает от его износа и позже приходит к старению и утрате умственных способностей.