Основная статья
:
Цветное зрение
Основная статья
:
Теория трёхкомпонентного цветного зрения (версия Миг)
Рис.1. Мозг человека, вид сзади. Красным цветом обозначена
первичная зрительная кора V1
(поле Бродмана 17) ; оранжевым — поле 18; жёлтым — поле 19.[1] Рис.2. Мозг человека, вид слева. Вверху: латеральная поверхность, внизу: медиальная поверхность. Оранжевым цветом обозначено поле Бродмана 17 (первичная, или стриарная, зрительная кора)[2] Рис.3. Дорсальный (зелёный цвет) и вентральный (сиреневый цвет) зрительные пути, берущие начало в первичной зрительной коре.[3]
Зрительная кора
(англ. visual cortex) является частью коры больших полушарий головного мозга, отвечающая за обработку зрительной информации. Главным образом она сосредоточена в затылочной доле каждого из полушарий головного мозга[4].
Оппонентно выделенные самые яркие сигналы видимых лучей света S,M,L — КЗС (не в цвете), сфокусированных предметных точек на экстерорецепторы (версия Миг) колбочки сетчатки глаза (рецепторный уровень), по зрительным нервам пересылаются сюда, в зрительную кору. Здесь формируется бинокулярное (стерео) цветное оптическое изображение (нейронный уровень). Впервые, субъективно мы ощущаем цвет (версия Миг), который является лично нашим. (При определении цвета методом колориметрии цвет оценивается данными среднестатистического наблюдателя большой группы здоровых людей)
Понятие зрительная кора
включает
первичную зрительную кору
(также называемую
стриарной корой
или
зрительной зоной V1
) и экстрастриарную зрительную кору — зоны
V2
,
V3
,
V4
, и
V5
. (См. о зонах
V2
,
V3
,
V4
, и
V5
в статье Зрительная кора)
Первичная зрительная кора анатомически эквивалентна полю Бродмана 17
, или BA17. Экстрастриарная зрительная кора включает поля Бродмана 18 и [4].
Зрительная кора присутствует в каждом из полушарий головного мозга. Области зрительной коры левого полушария получают сигналы от правой половины зрительного поля, правого полушария — получают сигналы от левой половины.
В дальнейшем в статье будет говориться об особенностях зрительной коры приматов (в основном, человека).[5]
Введение[править]
Рис.4,Схема цветного зрения с точки зрения трёхкомпонентной теории
Зрительные отделы головного мозга
— восприятие цвета и света, получение оптического изображения в коре головного мозга — второй, окончательный этап работы зрительной системы образования оптического видения в зрительных отделах головного мозга (см. рис.3,4).
Даже на начальном этапе визуального восприятия света и цвета в визуальной системе, в пределах сетчатки, проходя через начальные цветные механизмы «противника».
Рис.3a. Оптические тракты после встречи сигналов от правого и левого глаза в слои коленчатого тела
Известно, что механизмы противника обращаются к противостоящему цветовому эффекту красно-зелёных, синих-жёлтых и чёрно-белых цветов. (См.Теория оппонентного цветного зрения (версия Миг)). При этом визуальная информация возвращается назад через зрительный нерв к оптическому перекрёстку, где два оптических нерва встречаются и информация от временных (контралатеральных) визуальных полевых перекрёстков до противоположной стороны мозга. После оптического перекрёстка зрительные тракты нервного волокна упоминаются как оптические тракты, которые входят в таламус en:Thalamus через синапс в боковом латеральном коленчатом теле (ЛКТ). ЛКТ является отдельным отделом головного мозга из шести слоёв: два магноцеллюлярных (большая клетка) бесцветные слои (М. клеток) и четырёх parvocellular (маленькая клетка) цветных слоёв (P клетки). В пределах слоев P-клетки ЛКТ есть два цветных типа противника: красного против зелёного и синего против жёлтого (зелёного/красного)
.
После синапсиса в ЛКТ, визуальные тракты продвигается назад к первичной зрительной коре
(ПВК-V1), расположенной позади мозга в пределах затылочной доле. В пределах V1 слоя наружного коленчатого тела есть отличная полоса (бороздчатость). Это также упоминается «как полосатая кора», с другими корковыми визуальными областями, упомянутыми все вместе как «extrastriate кора». На данном этапе цветная обработка становится намного более сложной.
Активирующая ретикулярная система
Диффузную нервную сесть составляют неспецифические ядра. Находится эта система в медиальных отделах таламуса. Он является передним отделом активирующей ретикулярной системы, регулирующей возбудимость коры. Разнообразные сенсорные сигналы способны активировать данную систему. Сенсорные сигналы могут быть как зрительными, так и обонятельными, соматосенсорными, вестибулярными, слуховыми.
Активизирующая ретикулярная система представляет собой канал, который передает к поверхностному слою коры данные сигналов через расположенные в таламусе неспецифические ядра. Возбуждение АРС необходимо для того, чтобы человек был способен поддерживать состояние бодрствования. Если в данной системе возникают нарушения, то могут наблюдаться коматозные сноподобные состояния.
Первичная зрительная кора (VI)[править]
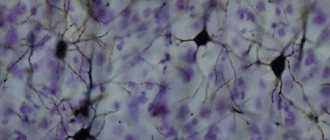
Рис.4. Мозг человека. Красным цветом обозначена первичная зрительная кора (зрительная зона V1
) Рис.5.Микрофотография показывает зрительную кору (розовый цвет). В pia mater и паукообразные в том числе кровеносные сосуды видны в верхней части изображения. Подкорковое белое вещество (синий) — это видно в нижней части изображения. ОН-LFB пятно..
Первичная зрительная кора
— наиболее изученная зрительная зона мозга. Исследования показали, что у млекопитающих она занимает задний полюс затылочной доли каждого полушария (эти доли ответственны за обработку зрительных стимулов). Это наиболее просто устроенная[6] и филогенетически более «старая» из кортикальных зон, связанных со зрением. Она приспособлена для обработки информации о статических и движущихся объектах, в особенности, для распознавания простых образов.
Составная часть функциональной архитектуры коры больших полушарий головного мозга — первичная зрительная кора — практически полностью соответствует анатомически определяемой стриарной коре
. Название последней восходит к латинскому «полоса, полоска» (лат. stria) и во многом обусловлено тем, что здесь отчётливо видна невооружённым глазом
полоска Дженнари[en]
(наружная полоска Байярже), образованная конечными отделами покрытых миелиновой оболочкой аксонов, отходящих от нейронов латерального коленчатого тела и заканчивающихся в
IV
слое серого вещества.
Первичная зрительная кора подразделяется на шесть функционально различающихся горизонтальных цитоархитектонических слоёв
(см. рис.К), обозначаемых римскими цифрами от
I
до
VI
[4][7].
Слой IV
(
внутренний зернистый слой
[7]), к которому подходит наибольшее количество афферентных волокон, идущих от латеральных коленчатых тел (ЛКТ), в свою очередь, подразделяется на четыре подслоя, обозначаемых
IVA
,
IVB
,
IVCα
и
IVCβ
. Нервные клетки подслоя
IVCα
, в основном, получают сигналы, идущие от нейронов магноцеллюлярных («крупноклеточных», вентральных) слоёв ЛКТ[8] («
магноцеллюлярный зрительный путь
»), подслоя
IVCβ
— от нейронов парвоцеллюлярных («мелкоклеточных», дорсальных) слоёв ЛКТ[8] («
парвоцеллюлярный зрительный путь
»).
Полагают, что среднее число нейронов первичной зрительной коры взрослого человека составляет около 140 миллионов в каждом полушарии[9].
Функция[править]
Рис.К. Полоса 6 — первичная зрительная кора (также называемую стриарной корой или зрительной зоной V1. Схема диаграммы P-клеткок нейронв, расположенных в пределах parvocellular слоев коленчатого ядра (LGN) таламуса
Первичная зрительная кора (V1) имеет очень четкие карты пространственной информации при зрении. Например, у людей верхняя половина области calcarine («шпоры») трещины отвечает на поступающие зрительные сигналы сильно. От нижней половины поля зрения области calcarine поток идёт в верхнюю половину поля зрения. Концептуально, это (retinotopic) или это отображение визуальной информации от сетчатки, нейронов, особенно визуального потока нейронов. Так происходит картирование — трансформация визуального оптического изображения от сетчатки в зону V1.
Соответствие данного расположения в зоне V1 и в субъективнои поле зрения — это соотносится очень точно: даже слепые пятна сетчатки сопоставляются с зоной данными в V1. С точки зрения эволюции, эта переалресация очень просто происходит у самых животных, которые обладают зоной V1. У животных и человека с fovea (центра макулы — жёлтого пятна) в сетчатке, большая часть зоны V1 сопоставляется с небольшой Центральной частью поля зрения. Явление, известное как корковые увеличения. Возможно, с целью точного пространственного кодирования, нейроны в V1, имеют наименьшие рецептивное поле размеров любой зрительной коры или микроскопические участки.
Настройка свойств нейронов зоны V1 (реакции нейронов) отличаются значительно с течением времени. В начале времени (40 мс и далее) время настройки отдельных нейронов V1 имеют сильные (тюнинг) характеристики воздействия небольшого набора стимулов. То есть ответы неёронов могут различаться небольшими изменениям в зрительной ориентации пространственных частот и цвета. Более того, отдельные нейроны человека и животных зоны V1 бинокулярного зрения у глазной системы, а именно : настройка одного из двух глаз. В зоне V1 и первичной сенсорной коре головного мозга в целом, нейроны с аналогичными настройки свойств имеют тенденцию объединяться в виде корковых столбцов. Дэвид Hubel и Торстен Визель предложены классические «кубики льда» — модель организации корковых столбцов для настройки двух свойств: глазных доминирований и ориентации. Однако эта модель не может вместить цвет, пространственную частоту и много других возможностей, которые настраивают нейроны [цитата]. Точная организация всех этих корковых столбцов в зоне V1 остается горячей темой настоящего исследования.
Текущий консенсус таков, что кажется, ответы работы нейронов зоны V1 состоят из плиточной структуры, которые представляют селективные пространственно-временные фильтры. Функционирование зоны V1 в пространственной области можно рассматривать как аналог множества пространственно-местных — комплекс Преобразования Фурье или ,точнее, трансформирование Габора. Теоретически эти фильтры вместе могут осуществлять обработку нейронов пространственной частоты, ориентацию, движение, направление, скорость (временной частоты), и многие другие пространственно-временные характеристики. Требуются эксперименты нейронов для обоснования этих теории, но постановку новых вопросв.
В более позднее время (после 100 ms) воздействия на нейроны зоны V1 они также чувствительны к более глобальной организациии сцены (Lamme & Roelfsema, 2000). Эти параметры ответа, вероятно, обусловлены повторяющейся обработкой (при влиянии высокого уровня областей коры головного мозга на нижний ярус областей коры головного мозга) и горизонтальными связями от пирамидных нейронов (хьюп et al. 1998). В то время как прямые соединения, в основном, в процессе работы, обратной связи, в основном — модуляторные с их последствиями (Angelucci et al., 2003; хьюп et al., 2001). Опыт показывает, что обратная связь, происходящих в более высшем уровне, в таких областях, как V4 ОН или MT, с более крупных и сложных рецептивных полей, может изменить и форму ответов зоны V1, учета контекстных или экстра-классических рецептивных полей эффекта (Guo et al., 2007; Huang et al., 2007; Sillito et al., 2006).
Визуальная информация передана зоне V1 не закодирована в терминах пространственной (или оптический) съемки, но, скорее это — локальный контраст. Например, для изображения, состоящего наполовину со стороной черного и половины стороны с белым цветом, разрыв строки между черным и белым представляет сильные местные контрасты и кодируется, и в то же время в виде нескольких нейронов кода информация о яркости (черный или белый per se). В качестве информации дальнейшей ретрансляции в последующие зрительные зоны, в ней закодированы также все нелокальные частоты, фазы сигналов. Главное, что на таких ранних этапах корковой визуальной обработки, пространственное расположение визуальной информации хорошо сохранилось на фоне локального контраста кодирования.[10]
Принцип оппонентности в отборе цветовых сигналов[править]
Основная статья
:
Цветное зрение — дифференцированное восприятие основных лучей сфокусированных предметных точек (версия Миг)
Основная статья
:
Трихроматизм и принцип оппонентности (версия Миг)
Основная статья
:
Теория оппонентного цветного зрения (версия Миг)
Схема трихроматизма с оппонентным отбором цвета Принципиальная схема трёхкомпонентрого цветного зрения человека, приматов с оппонентным отбором самых ярких основных лучей света КЗС с последующей передачей цветовых сигналов S,M,L в мозг
Принцип оппонентности отбора основных световых сигналов S,M,L — КЗС
— теория, которая определяет способ, которым сетчатка (версия Миг) человеческого глаза позволяет зрительной системе на нейронном уровне в коре голоаного мозга ощущать цвет (версия Миг). Это происходит на уровне восприятия видимых лучей (рецепторном) тремя разновидностями колбочек RGB (S,M.,L), на базе оппонентного отбора цвета (белый-чёрный, красный-зелёный, синий-жёлтый) с последущей передачей сигналов в мозг.[11][12]
Оптические изображения в мозгу и в фотографии[править]
Оптическое изображение в мозгу[править]
На основании вышеизложенного видно, что оптическое изображение (или предметные точки) на фокальной поверхности — сетчатке (биологическом фотосенсоре) как и в фотографии воспринято клетками, состоящими из определённого количества фотодатчиков (пикселей), например, колбочек, чувствительных к основным спектральным лучам, например, к красному, зелёнрму, синему (RGB). Сигналы фотодатчиков или фоторецепторов колбочек (их количество около 6 млн.) по строго связанной биологической системе передачи их при помощи синапсов по нерным каналам, которых насчитано примерно около 1,2 млн., передаюрся в головной мозг. Возникае вопрос, как это 6 млн. сигналов, трансдукцируемых синими, зелёными, красными колбочками каждого блока или от 2млн. клеток могут передаться по 1,2млн. каналов? При этом следует учесть работу экстерорецепторов (фотодатчиков) ганглиозного слоя сетчатки ipRGC, синапсически связанных прямой и обратной связью с колбочками, палочками и с головным мозгом, содержащими фотопигмент меланопсин, которые способны подавлять или усиливать фототрансдукцию биосигналов палочек и колбочек.[цитата, необходимая].
На начальном этапе визуального восприятия света и цвета (в пределах сетчатки) восприятие цвета начинается на раннем уровне в визуальной системе — уже в пределах сетчатки, проходя через начальные цветные механизмы «противника» — оппоннтного отбора наиболее ярких сигналов.
После синапсиса в LGN, визуальные тракты продвигается назад к первичной визуальной коре
(ПВК-V1)en:Visual_cortex, расположенной позади мозга в пределах затылочного лепестка en:Occipital_lobe. В пределах V1 слоя наружного коленчатого тела есть отличная полоса (бороздчатость). Это также упоминается «как полосатая кора» с другими корковыми визуальными областями, упомянутыми все вместе как «extrastriate кора». На данном этапе цветная обработка становится намного более сложной.
В итоге, созданный природой биологический АЦП (на уровне сетчатки и головного мозга) — уникальная биологическая система преобразования и получения оптического изобраежия (цветного и серого) в мозгу (в том числе и стерео). Достижения в области фотографии цветной, стерео ещё далеки от совершенства этих визуальных биологических систем, созданных природой, обладая которыми мы каждый день визуально наслаждаемся окружающим нас красочным миром.
Оптическое изображение в фотографии[править]
Основная статья
:
Фильтр Байера
Промежуточный мозг
Между больших полушарий находится промежуточный мозг, который подразделяется на две части: таламус и гипоталамус. Помимо них, выделяют еще эпиталамус, к которому примыкают эпифиз и гипофиз — эндокринные железы. Таламус — это «информационная воронка», которая фильтрует сигналы и пропускает их в кору больших полушарий: если бы все информационные потоки проходили в кору, она не смогла бы эффективно функционировать. Блокирование сигналов осуществляется с помощью тормозных нейронов. Структуры таламуса соответствуют разным центрам коры больших полушарий: передние ядра отвечают за передачу информации в центры эмоций и памяти, вентральные боковые связаны с двигательным контролем, вентробазальный комплекс работает с информацией о чувствительности тела, а над ним находятся слуховые и зрительные центры. Медиальные ядра таламуса связаны с центрами сна и бодрствования, а также с вкусовыми и болевыми сигналами и вестибулярной чувствительностью.
Гипоталамус занимается нейроэндокринной регуляцией и управляет деятельностью различных внутренних органов. Кроме того, в нем находятся важнейшие центры биологических потребностей: голода и жажды, полового и родительского поведения, страха и агрессии. Нейроны гипоталамуса оценивают концентрацию основных гормонов в крови. Гипоталамус тесно взаимодействует с гипофизом — железой, которая вырабатывает тиреотропный гормон и тем самым регулирует активность щитовидной железы. Команды гипофизу отдаются с помощью рилизинг-гормонов, которые вырабатывает гипоталамус. Также в гипоталамусе вырабатываются окситоцин и вазопрессин — гормоны, соответственно отвечающие за сокращение матки при родах и молочных желез при кормлении ребенка и потребность организма в жидкости. Эпифиз (или шишковидное тело) влияет на половое развитие и сексуальное поведение и вырабатывает мелатонин, который участвует в синхронизации циркадных ритмов.
О работе таламуса и гипоталамуса
Примечания
- Butler, Ann B. (2001 ). «Chordate Evolution and the Origin of Craniates: An Old Brain in a New Head». The Anatomical Record 261: 111—125.
- Головной мозг земноводных
- Brain Facts and Figures (англ.). Проверено 8 июня 2012. Архивировано 22 июня 2012 года.
- Heike Le Ker.
Neuronen-Nachschub: Neue Nervenzellen wachsen im menschlichen Gehirn nach (нем.).
Spiegel Online
(15. Februar 2007). Проверено 9 января 2020. - Нервные клетки человеческого мозга все-таки восстанавливаются, утверждают шведские ученые. NEWSru.com
(16 февраля 2007). Проверено 9 января 2020. - Евгения Самохина.
«Прожигатель» энергии // Наука и жизнь : журнал. — М., 2020. — № 4. — С. 22—25. — ISSN 0028-1263.
Содержание
- 1 Головной мозг как орган позвоночных
- 2 Ткани мозга
- 3 Клетки мозга
- 4 Кровоснабжение
- 5 Функции
- 6 Отделы мозга
- 7 Пластичность
- 8 Эмбриональное развитие
- 9 Методы исследования 9.1 Абляции
- 9.2 Транскраниальная магнитная стимуляция
- 9.3 Электрофизиология
- 9.4 Электрическая стимуляция
- 9.5 Другие методики
Эмбриональное развитие
Мозг четырёхнедельного эмбриона
Эмбриональное развитие мозга является одним из ключей к пониманию его строения и функций.
Головной мозг развивается из ростральной части нервной трубки. Бо́льшая часть головного мозга (95 %) является производной крыловидной пластинки.
Эмбриогенез мозга проходит через несколько стадий.
- Стадия трёх мозговых пузырей — у человека в начале четвёртой недели внутриутробного развития ростральный конец нервной трубки формирует три пузыря: Prosencephalon (передний мозг), Mesencephalon (средний мозг), Rhombencephalon (ромбовидный мозг, или первичный задний мозг).
- Стадия пяти мозговых пузырей — у человека в начале девятой недели внутриутробного развития Prosencephalon окончательно делится на Telencephalon (конечный мозг) и Diencephalon (промежуточный мозг), Mesencephalon сохраняется, а Rhombencephalon делится на Metencephalon (задний мозг) и Myelencephalon (продолговатый мозг).
В процессе формирования второй стадии (с третьей по седьмую недели развития) головной мозг человека приобретает три изгиба: среднемозговой, шейный и мостовой. Сначала одновременно и в одном направлении формируются среднемозговой и мостовый изгибы, потом — и в противоположном направлении — шейный. В итоге линейный мозг зигзагообразно «складывается».
При развитии мозга человека можно отметить определённое сходство филогенеза и онтогенеза. В процессе эволюции животного мира первым сформировался конечный мозг, а затем — средний мозг. Передний мозг является эволюционно более новым образованием головного мозга. Также и во внутриутробном развитии ребёнка сначала формируется задний мозг как самая эволюционно древняя часть мозга, а затем — средний мозг и потом — передний мозг. После рождения с младенческого возраста до совершеннолетия происходит организационное усложнение нейронных связей в мозге.
Двигательная зона
Поговорим о двигательной зоне отдельно. Следует отметить, что эта зона коры никак не соотносится с долями, рассмотренными выше. Она является частью коры, содержащей прямые связи с мотонейронами в спинном мозге. Такое название носят нейроны, непосредственно управляющие деятельностью мышц тела.
Основная двигательная зона коры больших полушарий располагается в извилине, которая называется прецентральной. Эта извилина представляет собой зеркальное отображение сенсорной зоны по многим аспектам. Между ними имеется контралатеральная иннервация. Если сказать иными совами, то иннервация направлена на мышцы, которые расположены на другой стороне тела. Исключение – лицевая область, для которой характерен контроль мышц двусторонний, расположенных на челюсти, нижней части лица.
Немного ниже основной двигательной зоны расположена дополнительная зона. Ученые полагают, что она имеет независимые функции, которые связаны с процессом вывода двигательных импульсов. Дополнительная двигательная зона также изучалась специалистами. Эксперименты, которые ставились над животными, показывают, что стимуляция этой зоны провоцирует возникновение двигательных реакций.
Особенностью является то, что подобные реакции возникают даже в том случае, если основная двигательная зона была изолирована или разрушена полностью. Она также вовлечена в планирование движений и в мотивацию речи в полушарии, которое является доминантным. Ученые полагают, что при повреждении дополнительной двигательной может возникнуть динамическая афазия. Рефлексы головного мозга страдают.