Что это такое
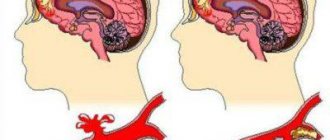
Ирритация коры головного мозга – патологическое состояние, проявляющиеся в виде спонтанного образования очага раздражения и возбуждения в определенной области коры головного мозга. Симптомы ирритации определяются локализацией патологического состояния.
Следует различать нормальную ирритацию – раздражение нервных волокон в ответ на внешний стимул и формирование адекватного ответа. Например, при воздействии яркого света на глаза сокращается зрачок (уменьшает поступление фотонов) в результате ирритации зрительного нерва. Патологическая же ирритация – это спонтанное раздражение, не имеющее очевидного происхождения и приводящее к ухудшению качества жизни больного.
Ирритация не входит в список самостоятельных заболеваний, ее нет в Международной Классификации Болезней 10-го пересмотра. Раздражение коры головного мозга выступает как проявление основной патологии, например, опухоли подкорковых структур.
Ирритация бывает очаговой, когда раздражение присутствует в отдельном участке коры (в зрительной или лобной) и диффузной (раздражается вся кора).
Ирритация коры головного мозга также бывает:
- Бессимптомной – раздражение коры может не доходить до порогового уровня и не вызывать признаков заболевания.
- Симптоматической – раздражение входит в порог чувствительности и определяет клиническую картину.
Высшие корковые функции человека и их нарушения при локальных поражениях мозга
Высшие корковые функции человека и их нарушения при локальных поражениях мозга
Оглавление
До сих пор мы оставались в пределах характеристики строения и функциональной организации отдельных областей коры головного мозга, пользуясь в основном методами морфологического и физиологического анализа отдельных участков мозговой коры.
За последние десятилетия развилась, однако, новая область науки, позволяющая сделать первые шаги к тому, чтобы проникнуть в наиболее тонкие и интимные механизмы мозговой деятельности. Речь идет о том, что в результате развития экспериментальной техники стало возможным приступить к физиологическому анализу работы отдельных нейронов, раздражая их электрическим током или отводя потенциалы, возникающие в них при различных условиях, меняющих поведение животного.
1 Раздел написан О. С. Виноградовой.
Нет никаких сомнений, что этот новый раздел науки дает существенную информацию об интимных механизмах работы нервной ткани и со временем позволит пересмотреть многие из сложившихся к настоящему времени представлений о функциональной организации отдельных разделов коры головного мозга.
Следует с самого начала отметить, что почти все имеющиеся данные о работе нейронов мозговой коры получены на кроликах или кошках и лишь небольшая часть исследований проведена на обезьянах. Это означает, что эти данные имеют лишь самый первоначальный характер, и переносить их на человека можно лишь с очень большой осторожностью. Однако, несмотря на это, данные, полученные при исследовании на нейронном уровне, уже сейчас дают существенный материал, который может служить для выяснения общих принципов функциональной организации корковых центров и основных тенденций их развития.
Зрительная кора
Тщательные исследования первичной зоны зрительной коры, проведенные в лаборатории Юнга (1958, 1961), показали наличие разных типов нейронов, обнаруживаемых при применении диффузного света. Юнг выделил 5 основных типов: А — нейроны, не отвечающие на свет; В — отвечающие пачкой импульсов на включение света; С — отвечающие торможением на включение света; D — отвечающие на выключение света; E — отвечающие на включение и выключение света. Нейроны типа А, по данным Юнга, составляют примерно 50% всех нейронов зрительной коры. По данным других авторов, число нейронов, не отвечающих на диффузный засвет, еще больше. Как было показано Хьюбелом (1962), специальный подбор зрительных сигналов позволяет получить ответы со всех зрительных нейронов. Однако для этого необходимо использовать оформленные, структурированные зрительные сигналы. Хьюбел показал, что это зависит от особой организации рецептивных полей клеток зрительной области. Он выделяет, прежде всего, клетки с так называемыми «простыми рецептивными полями ». В отличие от нейронов латерального коленчатого тела с их концентрическими полями эти клетки имеют вытянутые активные зоны, зависящие от величины, формы, положения и направления движения раздражителя. Каждая такая активная зона окружена областью, оказывающей антагонистическое, тормозное’ влияние на реакции, вызываемые с центра данной области. Именно поэтому диффузный засвет, возбуждая одновременно активный центр и тормозную периферию рецептивного поля нейрона, препятствует его ответу. Кроме указанных нейронов Хьюбел обнаружил клетки с так называемыми «сложными рецептивными полями». Эти поля обширнее и в меньшей степени зависят от точной формы и размера стимула. Для некоторых нейронов активным раздражителем является край, контур, светлый с одной стороны и темный — с другой. Активные стимулы для нейронов такого типа можно в довольно широких пределах варьировать по величине и углу наклона. Однако общая форма и направление движения стимула сохраняют свое значение. По мере удаления от области п роекции макулы поля нейронов возрастают по величине. Хьюбел рассматривает эти нейроны как интеграторы, на которые конвергируют несколько клеток с «простыми полями», дающие «абстракцию направления и формы независимо от точного положения объекта в зрительном поле».
Таким образом, корковая проекция сетчатки (поле 17) по данным нейронных исследований уже не представляет собой простую мозаику топографических пунктов («point-by-point»), a является органом интеграции высокого порядка, специально приспособленным для предметного зрения.
Интересно, что при строго перпендикулярном к поверхности коры перемещении электрода обнаруживается, что все нейроны, лежащие друг под другом, имеют сходные рецептивные поля. Тем самым выдвигается принцип организации корковых зон анализаторов в «функциональные колонки»; с этим принципом мы встретимся и далее при рассмотрении организации других областей.
Описанными нейронами не исчерпывается функциональная мозаика зрительной области. Здесь обнаружены нейроны, отвечающие только на монохроматические источники света в узкой полосе спектра (60—100 м\х) (Андерсен, Бухманн, Леннокс, 1962). Эти нейроны, составляющие примерно 7з всех элементов, не отвечают на белый свет, и их ответ не зависит от яркости. Они выделяют качество света в широких пределах независимо от иных характеристик раздражителя. Наряду с этим Берне (1964) показал, что некоторые клетки не меняют активности при различных изменениях свойств сигнала, но отмечают все сдвиги в их интенсивности.
Многие элементы первичной зрительной области отвечают на вестибулярные раздражения (Юнг, 1962). В этих случаях наблюдается конвергенция зрительных и вестибулярных влияний на один и тот же нейрон, причем вестибулярные влияния могут в значительной степени модулировать ответы на зрительные сигналы. Поэтому, как указывает Юнг, здесь происходит не только прием дискретных форм зрительной информации, но и ее интеграция с вестибулярными и оптокинетическими раздражителями. Вместе с тем в первичной зрительной коре было найдено очень небольшое число (не более 5%) нейронов, обнаруживающих бинокулярную конвергенцию. В случае выявления конвергенции ответы возникали при локальном раздражении гомотопических участков сетчатки двух глаз. При этом обычно влияния были антагонистическими: например, возбуждение с контрлатерального глаза и торможение с гомолате-рального. Истинная суммация бинокулярных раздражений наблюдалась так редко, что большинство исследователей склоняется к выводу, что механизмы бинокулярного зрения обеспеч иваются, во всяком случае, не в первичной зрительной коре.
Наконец, следует указать еще один факт, наблюдавшийся многократно различными исследователями. Довольно большое число нейронов первичной зрительной области отвечает на звуковые, тактильные, обонятельные и болевые воздействия. Эти ответы более вариабельны, чем реакции на свет, имеют большие латентные периоды и часто конвергируют на нейроны, отвечающие на световые сигналы. Их функциональное значение не вполне ясно, можно только утверждать, что большинство из них не связано с ориентировочным рефлексом и вниманием, поскольку они сохраняют свою работу без тенденции к ослаблению при длительном применении сигнала.
Слуховая кора
В первичной слуховой зоне кошки на нейронном уровне отмечается наличие тонотопической организации, хотя данные разных авторов по этому вопросу не вполне согласуются между собой. Кацуки (1962) отмечает, что в передней части эктосильвиевой извилины располагаются преимущественно клетки, отвечающие на высокие тоны, в средней — на тоны средних частот и в задней — на низкие тоны. Другие авторы (Эванс, Росс, Уайтфилд, 1965) считают на основании своих данных, что клетки, отвечающие на высокие тоны (больше 10 кгц), действительно находятся в передней части зоны Al, но клетки с ответами на более низкие звуки равномерно распределены по всей слуховой коре, так что подлинная тонотопическая организация отсутствует. Во всяком случае несомненно одно: на клеточном уровне такая организация оказывается значительно более сложной, поскольку нейроны слуховой коры характеризуются не просто определенными звуковысотными параметрами, а значительно более сложными функциональными характеристиками, аналогично тому, как это было показано выше для нейронов зрительной коры.
Предлагавшиеся классификации слуховых нейронов весьма сложны и разнообразны (Оониши, Кацуки, 19G5), поэтому мы ограничимся описанием некоторых типов ответов, наблюдавшихся в основном в слуховой коре кошки.
Большинство нейронов в зоне Al отвечает на применяемые звуки (краткие или длительные) краткими фазическими реакциями на включение. Реже наблюдаются нейроны с тоническим типом ответа, реагирующие длительным повышением или снижением уровня активности в течение всего времени действия звука. Ряд нейронов имеет узкую оптимальную частоту1, не отвечает на другие сопредельные частоты или отвечает на них значительно слабее и при значительно более высоких порогах. Галамбосом и другими был, например, описан в слуховой коре кошки нейрон, отвечавший на тон 235 гц (но не на 234 или 236 гц), что говорит о чрезвычайной тонкости слухового анализа. Однако наиболее типичным для слуховой коры являются клетки с очень широкими полосами оптимальных частот, значительно превосходящими по широте параметры, характерные для клеток медиального коленчатого тела. Это, как считает Кацуки, говорит о том, что полный звуковысотный анализ, по всей вероятности, заканчивается на таламическом уровне, а кортикальные нейроны проекционной области служат уже интеграторами, на которые конвергируют несколько элементов медиального коленчатого тела. Это подтверждается и тем фактом, что в коре обнаруживаются клетки с несколькими пиками на кривой реактивности, т. е. с несколькими оптимальными частотами в разных участках звукочастотной шкалы, что никогда не наблюдается на низших уровнях слухового анализатора. Существенно, что в кортикальных нейронах Кацуки не наблюдал характерной для всей слуховой системы зависимости параметров ответа от интенсивности звука, описываемой S-образной кривой. Только очень резкие и сильные изменения звука могут отражаться на реакциях корковых нейронов (обычно подавляя их), изменения же интенсивности в пределах среднего диапазона не отражаются на ответах нейронов. Следовательно, функция интенсивности кодируется клетками низших уровней, а корковые нейроны имеют дело с выделением более информативных и тонких характеристик звука.
Реакции нейронов слуховой коры значительно усиливаются’при одновременном нанесении двух различных звуков. При этом особенно отчетливые ответы наблюдаются, если звуки находятся в гармоническом отношении (т. е. их частоты относятся как 1 :2, 1 :3 и т. д.) и образуют биения. В этих случаях краткие фазические ответы на включение тока превращаются в длительные ритмические реакции. Это явление Кацуки связывает с восприятием на уровне коры сложных тембровых характеристик звука. 10% нейронов слуховой коры отвечает только на частотно-модулированные тоны. При этом многие из них имеют частотную ориентацию, т. е. отвечают только при повышении или только при снижении частоты тона.
1 «Оптимальная частота» — понятие, соответствующее «рецептивному полю» в зрительном и соместетическом анализаторах — частота, вызывающая максимальный ответ нейрона при наименьшем пороге.
Конечно, в области слуха еще более трудно делать выводы из данных, полученных на животных, но описанные факты позволяют считать, что отдельные нервные элементы, которые уже у кошки могут выделять столь многообразные и сложные характеристики звука, у человека могут дифференцироваться далее для анализа отдельных фонем и сложных акустических параметров речевых звуков. Галамбосом и др. (1958) описана в первичной слуховой коре кошки группа нейронов (около 10%), отвечавших только на сложные значимые звуки (зов, мяуканье, писк мыши). Правда, эти данные можно интерпретировать самым различным образом — не обязательно как результат специфической кортикальной интеграции, но как следствие конвергенции влияний от активирующих и «эмоциональных» систем (неспецифический таламус, лимбические образования) на данные нейроны.
Основным нейронам слуховой коры с их достаточно широкими частотными характеристиками, по данным Кацуки, присуща анатомическая, организация в вертикальные столбики с одинаковыми функциональными свойствами. Здесь также обнаружены отдельные элементы, отвечающие на раздражители незвуковых модальностей.
Соматосенсорная кора
Задне-центральная кора в области, соответствующей соматотопиче-ской проекции соматической чувствительности, исследовалась на кошках и обезьянах Маунткаслом с сотр. (1957, 1959, 1966). Эту область в основном занимают элементы с локальными рецептивными полями и достаточно четким соматотопическим расположением. Рецептивные поля иногда могут быть очень ограниченными (2—8 см2 на периферии), однако, как правило, рецептивные поля в коре в 15—100 раз больше полей соматических афферентных нейронов первого порядка. Подавляющее большинство элементов этой корковой области отвечает только на раздражение контралатеральной половины тела. Возбуждающее рецептивное поле таких нейронов окружено зоной, часто довольно сложной и неправильной конфигурации, с которой можно вызвать подавление ответа клетки на стимуляцию центра рецептивного поля. В области, лежащей кпереди от соматосенсорной зоны, на переходе к моторной коре передней части полушария, постепенно нарастает число нейронов с огромными диффузными рецептивными полями — о твет этих клеток часто можно вызвать со всех четырех конечностей или со всей половины тела, причем гомолатеральные и контралатеральные раздражения конвергируют на один нейрон. Латентные периоды здесь больше, а сами поля лабильнее, чем в ядре соматического анализатора. Эти нейроны могут рассматриваться как интеграторы полей нейронов ядерной зоны.
Характеристики соматических нейронов не исчерпываются, однако, их топическим представительством. Некоторые клетки отвечают только на раздражители определенных субмодальностей: на прикосновение к коже, потягивание за волоски, давление на глубокие ткани, смещение конечности в суставе. При этом раздражения разных модальностей могут находиться в реципрокных отношениях, и ответ на одну субмодальность (прикосновение) может подавляться иным раздражением того же рецептивного поля (давление). Маунткасл обнаружил в опытах на обезьянах преимущественное расположение основных субмодальностей по цитоархитектоническим полям (поле 3 — кожная чувствительность; поле 2 — глубокая чувствительность; поле 1 — переход между ними).
Однако наряду с этим существуют и нейроны, на которые конвергируют различные виды чувствительности, так что одна клетка может отвечать на сгибание пальцев, давление на кожу и потягивание за волоски.
Ответы клеток могут быть фазическими и тоническими. Например, при повороте конечности в суставе одни клетки отвечают короткой пачкой импульсов в момент поворота, а другие сохраняют повышенный уровень активности в течение всего времени сохранения конечностью измененного положения. Таким образом, одни элементы дают информацию о движении, а другие — о поддержании позы. Но даже в элементах с широкой конвергенцией сигналов, по мнению ряда исследователей, различные суб-модальноети или точки поверхности могут кодироваться в распределении разрядов клетки (pattern), в результате чего специфичность информации частично сохраняется, несмотря на ее обширную конвергенцию.
Принцип вертикальной организации коры был впервые показан Маунткаслом именно на примере соматосензорной области. Он показал значительное сходство рецептивных полей в колонках нейронов, регистрируемых при прохождении коры микроэлектродом, ориентированным перпендикулярно к ее поверхности. Такие функциональные колонки показаны им не только для топических полей, но и для характерных субмодальностей. На этом основании Маунткасл рассматривает вертикальные колонки как интегральные функциональные единицы коры, осуществляющие дискретный анализ и синтез соответствующих сигналов.
Вторичные зоны анализаторов
Данные относительно непервичных областей корковых анализаторов значительно более скудны и менее определенны, чем изложенные выше данные о первичных проекционных зонах. Это вполне понятно, так как у некоторых экспериментальных животных (кролик) сами эти области плохо выражены и по-разному определяются разными экспериментаторами-физиологами. С другой стороны, как мы показали выше, исследование самих первичных зон требует от экспериментатора тщательного подбора весьма сложных и своеобразных стимулов. При изучении непроекционных зон анализаторов поиски адекватных сигналов становятся еще более трудными, и различные исследования нейронов этих областей оставляют впечатление, что данные в известной мере упрощены за счет неадекватности примененных сигналов. В связи с этим мы ограничимся только перечислением лишь общих отличительных черт вторичных областей, выявившихся в исследованиях разных авторов.
В передней части латеральной, супрасильвиевой, эктосильвиевой извилинах, по данным регистрации суммарных вызванных потенциалов, в равной степени регистрируются вторичные ответы на световые, звуковые и кожные раздражения. При микро-электродных исследованиях в этих областях выявляется высокий процент нейронов с широкой мульти-сензорной конвергенцией. Обычно процент отвечающих нейронов здесь ниже, чем в проекционных областях, но большинство нейронов отвечает на 2—3 модальности. Вместе с тем нейронные иследования показывают отсутствие той однозначности, которая выступает при записи вызванных потенциалов. Например, в передней латеральной извилине кошки нейроны предпочтительно отвечают на звуковые сигналы и в меньшей степени— на световые и тактильные раздражители, а в супрасильвиевой извилине преобладают ответы на световые сигналы и т. д. Таким образом, каждая из областей имеет свой набо,р сензорных интеграции и — соответственно — специфические функции (Дубнер, 1966).
Наиболее глубокое исследование специфических сензорных функций непервичных областей зрительной области (поле 18 и 19 Бродмана) недавно проведено Хьюбелом и Визелом (1966). По их данным, эти области, как и первичная зона, имеют топографическую организацию; однако, возможно, что периферические области сетчатки здесь вообще не представлены. В поле 18 и 19 обнаружено прогрессивное усложнение рецептивных полей нейронов, причем изменение функциональных характеристик точно совпадает с цитоархитектоническими границами полей. В поле 18 почти отсутствуют простые поля (см. выше) и 90% составляют клетки со сложными полями. Здесь же появляются клетки с гиперсложными полями (5—10%). Гиперсложные клетки низшего порядка отвечают на зрительные комплексы («ограниченный движущийся край» и «дважды ограниченный движущийся край») ; несколько упрощая, можно сказать, что они отвечают на углы и прямоугольники, строго округленные по величине, ширине окружающего «ограничивающего края», ориентации и направлению движения. Гиперсложные клетки высшего порядка отвечают на две группы зрительных сигналов с ориентацией, различающейся на 90°. По своим свойствам их поля отчетливо характеризуются как результат синтеза нескольких сложных или гиперсложных полей низшего порядка. Такие элементы составляют более половины клеток 19 поля. Отдельные клетки этого поля имеют сложные поля, причем в вертикальных колонках часто обнаруживаются сложные и гиперсложные клетки с одинаковой ориентацией полей, образующие дискретные функциональные системы.
Таким образом, в поле 18 и 19 на нейронном уровне преобладает обобщенный прием сложной зрительной информации с выделением не контура (как в поле 17), а нарушений непрерывного контура (разрывы линий, кривизна, повороты, изменение направления движения). Кроме того, в 18 поле, по всей видимости, в значительной мере происходит бинокулярный синтез информации. Особой спецификой обладает вторичная соматическая зона, где наблюдается сохранение известной топической организации соматосензорного представительства, но в отличие от первичной зоны в этом представительстве обе половины тела (гомо-и контралатеральная) наложены друг на друга. Физиологический смысл этого факта не вполне ясен. Здесь же есть много нейронов с очень большими рецептивными полями, нередко они отвечают также на звуки (Маунткасл, 1962; Каррерас, 1963).
Нейроны вторичной слуховой области (А II) часто вообще не имеют оптимальной частоты и реагируют на звук независимо от его физических характеристик (Кацуки, 1962). Во вторичных областях слуховой коры значительно большее число нейронов имеет ответы тонического типа. Реакции возникают после более длительного латентного периода, но удерживаются в течение всего действия сигнала и нередко сохраняются на некоторое время после его прекращения, в то время как в первичной доминируют фазические ответы клеток на включение звукового сигнала.
Ответы нейронов вторичных областей значительно лабильнее, они вариативны и по рецептивным полям, и по воспроизведению структуры ответа (распределения зарядов во времени). Эти ответы в первую очередь подавляются при введении малых доз наркотиков, что говорит об их мультисинаптической природе (Каррерас, Андерсен, 1963).
Из изложенных данных следует, во всяком случае, что функции анализа элементарных физических характеристик сигналов присущи вторичным областям в значительно меньшей степени. Некоторые исследователи на этом основании приписывают мультимодальным клеткам вторичных областей роль диффузных активаторов, принимающих воздействия с неспецифических активирующих систем мозга и регулирующих «на месте» общий уровень активности мозга (включая организацию ориентировки и внимания). Однако, вероятно, что кажущаяся «упрощенность» суммарных реакций этих клеток является, как мы говорили выше, следствием примитивных, неадекватных форм воздействия. Анализ межнейронных связей и данные о структурной организации этих областей, приведенные в предыдущем разделе, позволяют считать, что нейроны вторичных областей являются истинными интеграторами, обобщающими в пределах одного или нескольких анализаторов первично проанализированную информацию.
Моторная кора
Данные о нейронах моторной коры (4—6 поле), по данным разных авторов, весьма однотипны. Ответы в этой области регистрируются обычно на значительной глубине (более 1000 μ от поверхности), в силу чего элементы, с которых производится запись, обычно отождествляются с большими пирамидами Беца в 5—6 слое. 3U всех клеток отвечают на соматическое раздражение более чем двух конечностей. Доминируют ответы на раздражение глубоких тканей, на растяжение и укалывание мышц, сгибание конечностей в суставах. Однако значительное число нейронов отвечает и на тактильную стимуляцию кожи. Сюда же конвергируют сигналы от вестибулярного аппарата и мозжечка. Все авторы без исключения отмечают широкую конвергенцию слуховых и зрительных возбуждений (Альб-Фессар, 1964; Бюзер, 1Θ61; Соколова, 1966). Активность ряда нейронов значительно учащается при осуществлении движения. В опытах на ненаркотизированных обезьянах было обнаружено, что в ряде случаев нейроны, отчетливо включающиеся в реакцию при активном движении конечности, не изменяют фоновой импульсации при пассивных перемещениях той же конечности (Эвартс, 1965). В других случаях наблюдается учащение активности, опережающее движение и совпадающее с общей реакцией активации в коре.
Все это позволяет рассматривать нейроны моторной коры как элементы истинного «конечного пути» или общую зону сензомоторной интеграции. Такой универсальной конвергенции не наблюдается ни в одной зоне задних отделов полушарий. Очевидно, конечные результаты анали-тико-синтетической работы задних отделов выводятся на моторные пирамиды для управления произвольными двигательными реакциями организма.
Данные о работе нейронов премоторных и фронтальных отделов мозга в настоящее время, к сожалению, отсутствуют.
Динамические характеристики кортикальных нейронов
Как подчеркивает большинство исследователей, изучавших нейроны первичных сензорных областей, их характеристики обладают строгим постоянством. Сигнал, вызывающий ответ нейрона проекционной области, обычно кодируется в определенной последовательности разрядов, меняющейся только при изменении тех или иных свойств сигнала. Строго постоянными являются и рецептивные поля нейронов. К сожалению, в настоящее время отсутствуют возможности регистрации ответов одного нейрона в течение длительных периодов времени (несколько дней), но регистрация в течение 8—10 часов в подавляющем большинстве случаев не обнаруживает изменения характеристик ответов нейрона. Поэтому предполагают, что указанные характеристики складываются на основании анатомических связей нейронов и в процессе функционирования на начальных этапах (Хьюбел и Визел, 1963), закрепляясь затем как функциональные единицы, образующие систему анализатора.
Однако кора всегда рассматривалась как основной аппарат обучения, внимания, памяти и индивидуального опыта. Поэтому естественно желание исследователей обнаружить на нейронах коры корреляты этих динамических процессов.
В ряде исследований было показано, что ответы нейронов меняют характеристики при стимуляции активирующей ретикулярной формации и неспецифического таламуса (Юнг, 1958; Фустер, 1951). При этом начинают отвечать ранее неактивные клетки, увеличивается вероятность ответов, а в нейронах зрительной коры возрастает критическая частота слития мельканий. Эти явления могут рассматриваться как аналоги механизмов внимания. Было показано также, что ответы слуховых нейронов увеличиваются при внимательном рассматривании источника звука. В зрительной коре кошки рассматривание мыши вызывает общее повышение частоты импульсации при одновременном снижении реакций нейронов на диффузную световую вспышку. В зрительной, слуховой, сомато-сензорной и моторной областях были обнаружены нейроны («детекторы новизны»), отвечающие только, если раздражитель является новым или необычным. При повторении сигнала ответы таких нейронов быстро ослабевают и полностью исчезают, но могут быть восстановлены изменением действующего сигнал а в каком-либо отношении (Хьюбел, Галамбос и др., 1959; Виноградова и Линдслей, 1963, Мурата и Камеда, 1963 и др.). Активность этих нейронов может (рассматриваться не только как аналог ориентировочного рефлекса или сензорного внимания, но и как коррелят фиксации следа раздражителя в системе памяти.
После начальных исследований Джаспера и сотр. (1958), показавшего сложное перераспределение активности в нейронах различных областей коры у обезьяны в процессе выработки двигательного условного рефлекса, в самые последние годы появилось много работ, воспроизводящих выработку условных связей на одиночных нейронах. При сочетании двух раздражителей, из которых лишь один («подкрепляющий») вызывает исходный ответ, удается получать «условный» ответ и на ранее неактивный раздражитель. Существенно, что появляющаяся при сочетании «условная» реакция отнюдь не воспроизводит ответ на «подкрепляющий» сигнал, что говорит о сложном характере формирующейся интеграции.
Хотя в настоящее время существуют лишь отдельные намеки, можно предполагать, что элемент динамических преобразований будет более выражен во вторичных и третичных образованиях коры. В первичных зонах эти процессы кажутся все же скорее исключением, чем правилом. Так, число «детекторов новизны» в зрительной и слуховой областях составляет всего 4—5%. Создается впечатление, что в первичных зонах функции строятся таким образом, чтобы обеспечить максимальную стабильность принимаемой и передаваемой информации, как «объективного» отражения внешних воздействий, в значительной мере не зависимого от отношения, интереса, значения и эмоциональной окраски сигнала. На основе этого стабильного отражения в дальнейшем, и территориально — в других областях, происходит вторичная обработка — отбор информации по индивидуальной значимости, фиксация в памяти и т. д. Возможно, что в некоторых отношениях такая обработка происходит в непервичных областях коры.
В настоящее время есть основания полагать, что в этих процессах принимает какое-то участие древнейшая область коры — гиппокамп. Исследование нейронов этой области обнаружило необычайно широкую конвергенцию сигналов разных сензорных модальностей. Однако все эти сигналы вызывают своеобразные тонические ответы нейронов только пока они обладают качеством новизны. Реакции быстро адаптируются и восстанавливаются при изменении любых параметров раздражителя. Нейроны с подобной динамикой ответов составляют около 80% всех реактивных нейронов гиппокампа, что заставляет предполагать существование особой роли гиппокампа в процессах различения известных и неизвестных сигналов, фиксации новых сигналов и подавления ответов на сигналы, уже Зафиксированные в прошлом опыте организма (Виноградова, 1964).
Таковы краткие данные о первых шагах в создании функциональной цитбархитектоники коры.
Причины
Патологическая ирритация коры головного мозга имеет такие причины:
- Воспалительные болезни нервной системы: нейросифилис, герпетический энцефалит, менингит.
- Осложнения основных заболеваний: малярия, краснуха, корь, менингококковый энцефалит.
- Нарушение кровообращения в мозгу: атеросклероз, транзиторная ишемическая атака, эмболия.
- Нарушение внутричерепного давления вследствие опухоли.
- Черепно-мозговые травмы: сотрясение, ушиб.
- Дислокационный синдром.
- Вредные привычки.
- Работа и проживание в загрязненных условия.
Общая информация
Менингиома представляет собой опухоль, чаще всего доброкачественную. Новообразование произрастает из клеток твердой мозговой оболочки и локализуется в черепной коробке человека, а именно на поверхности мозга или на основании черепа. Реже местом локализации менингиомы является желудочки мозга или костная ткань.
Данное новообразование характеризуется медленным ростом. Патология длительное время может протекать бессимптомно.
Симптомы
Признаки ирритации коры определяются локализацией раздражения. Симптомы непосредственно связанны с областью коры, где возникает очаговая спонтанная ирритация:
- Лобная зона. Сопровождается возникновением двигательных реакций. Сокращение мышцы зависит от места раздражения в прецентральной лобной извилине. После раздражения лобного участка могут появиться сложные двигательные паттерны: больной начнет завязывать шнурки в воздухе.
- Височная область. Появляются слуховые простые (акоазмы) и сложные галлюцинации, сопровождающиеся голосом комментирующего содержания.
- Затылочная зона. Сопровождается простыми (фотопсии) и сложными зрительными галлюцинациями. Фотопсии – это секундные галлюцинации: вспышки света, маленькое пятнышко. Сложные галлюцинации состоят из образов, содержание которых определяется внутренней психической жизнью пациента.
- Теменная зона– область общей чувствительности. Возникают покалывание, онемение, ползанье мурашек в разных частях тела. Раздражение в этой области также сопровождается извращенными ощущениями прикосновения, боли, жара или холода.
Диффузная ирритация коры сопровождается малыми (petit mal) и большими (grand mal) судорогами.
К малым припадкам относятся миоклонические судороги отдельных мышц. Сокращение мускула характеризуется ритмичностью и отсутствием осложнений. Petit mal также проявляется абсансами – кратковременными выключением сознания при сохранении тонуса мышц всего тела. После 20-30 секунд «выключения» больные приходят в себя и продолжают свою работу. Они не знают, что только что вышли из сознания.
Grand mal состоит из нескольких последовательных стадий:
- Предвестники. За день до обширных судорог люди ощущают недомогание, головную боль. Они плохо спят.
- Аура. За 30-40 минут больные жалуются на неопределенные по области боли в животе, руке или в сердце.
- Тоническая фаза. Человек теряет сознание, падает. Вся мускулатура тела сокращается одновременно и синхронно. Цвет кожи синеет, дыхание неравномерное. Длительность – не больше 60 секунд.
- Клоническая фаза. Все мышцы тела сокращаются неравномерно, несинхронно, хаотично: каждый мускул сокращается по отдельности. Длится 1-2 минуты.
В общем весь большой судорожный припадок длится до 3 минут. После последней фазы мышцы расслабляются, пациент уходит в глубокий сон. После пробуждения у него наблюдается дезориентация и ретроградная амнезия (не помнит, что было до припадка).
Конвекситальные менингиомы
Этот тип менингиом является наиболее часто встречаемым видом опухолевых новообразований. И диагностируется примерно в 50% от общего числа опухолей.
Наиболее часто встречающейся локализацией является такая, при которой конвекситальная менингиома головного мозга располагается под верхней крышкой коробки черепа (ее образуют правая и левая лобная, височная, теменная и затылочная кости). Такое расположение новообразования отмечается в 20% диагностированных случаев.
Диагностика заболевания
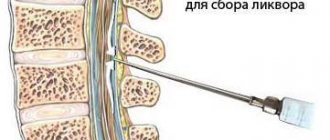
Для обследования патологии используется метод магнитно-резонансной томографии (МРТ), который дает подробную информацию о заболевании. Хорошо просматривается степень поражения патологии, размер опухолевого образования с разных ракурсов. Это позволяет собрать доброкачественный материал обследования патологии для назначения адекватного лечения.
При отсутствии возможности сделать МРТ существует другой вид диагностики – компьютерная томография. Он позволяет чётко рассмотреть новообразование на наличие кровоизлияний и отложения солей в ткани.
Информация, полученная с помощью МРТ и КТ, идентична. Однако во время МРТ человек не подвергается облучению. Чтобы отличить менингиому от других аналогичных опухолей используют метод МР-спектроскопии. При помощи магнитно-резонансных спектров можно изучить процессы метаболизма мозговых тканей.
Вспомогательным методом диагностики патологии является электроэнцефалография. Его применяют, когда нужно подтвёрдить, что эпилептические припадки вызваны опухолью. Гистологическое исследование – дополнительный метод, применяется после удаления опухоли. Он позволяет определить новообразование на злокачественность и понять, нужно ли продолжать лечение и назначать химиотерапию.
Последствия
Прогнозы при проведении лечебных мероприятий чаще всего благоприятные. Но стоит понимать, что и как любое заболевание, может иметь различный исход и конвекситальная менингиома. Последствия болезни чаще всего зависят от запущенности заболевания и места расположения опухоли в головном мозге человека. Возможны рецидивы болезни. Высоки риски рецидива опухоли, расположенной в теменной доле головного мозга.
Очень важно своевременно обратиться к врачу для получения специализированной помощи, поскольку опухоль ямки черепа может привести к полной потере обоняния, а новообразование в области турецкого седла может способствовать полной потере зрения.
Несмотря на то, что в большинстве случаев менингиома является доброкачественным образованием, данное заболевание может привести к нарушению работы слухового, зрительного и обонятельного анализаторов, к инвалидности. При нарушении работы жизненно важных центров возможен летальный исход.
Реабилитация после лечения
Длительность реабилитационного периода напрямую зависит от тяжести заболевания и места расположения опухоли в головном мозге человека. Помимо этого, на скорость восстановления пациента влияют те осложнения, которые были вызваны новообразованием.
В большинстве случаев при нарушении зрения и слуха, вызванных атрофией соответствующих нервов, восстановить нормальную работу анализаторов после проведения хирургического вмешательства невозможно.
При атипичных и злокачественных менингиомах после проведения курса лечения возможны рецидивы. Однако чаще всего при своевременном врачебном вмешательстве удается не только избавиться от новообразования, доставляющего дискомфорт, но и избежать негативных последствия и последующих осложнений.
Реабилитация пациента после курса лечения, направленного на борьбу с менингиомой, включает несколько важных пунктов:
- применение акупунктуры, которая способствует активизации нервных окончаний и восстанавливает чувствительность конечностей после паралича, вызванного опухолью;
- проведение медикаментозной терапии, которая поддерживает общий тонус организма и предупреждает возникновение рецидива;
- лечебная физкультура, благодаря которой человек может восстановить утраченные в результате болезни двигательные и другие функции организма.
Для быстрого и эффективного восстановления после курса терапии необходимо регулярно посещать лечащего врача и соблюдать все рекомендации.
Лечение
Конвекситальная менингиома лечится только хирургическим путем. С остальными видами менингиом можно бороться иначе.
В настоящее время существует огромное множество методов, позволяющих воздействовать на опухоль без проведения хирургического вмешательства. К таким методам относятся:
- стереотаксическая терапия;
- гамма-терапия;
- радиохирургия.
Вышеперечисленные способы лечения не только обладают высокой эффективностью, но и доступны широким слоям населения, поскольку данные методы недорогие.